Abstract
Tyrannosaurids were the most derived group of Tyrannosauroidea and are characterized by having two body plans: gracile, long-snouted and robust, deep-snouted skulls. Both groups lived sympatrically in central Asia. Here, we report a new deep-snouted tyrannosaurid, Asiatyrannus xui gen. et sp. nov., from the Upper Cretaceous of Ganzhou City, southeastern China, which has produced the large-bodied and long-snouted Qianzhousaurus. Based on histological analysis, the holotype of Asiatyrannus xui is not a somatically mature adult, but it already passed through the most rapid growth stages. Asiatyrannus is a small to medium-sized tyrannosaurine, with a skull length of 47.5 cm and an estimated total body length of 3.5–4 m; or around half the size of Qianzhousaurus and other large-bodied tyrannosaurines in similar growth stages. Asiatyrannus and Qianzhousaurus are sympatric tyrannosaurid genera in the Maastrichtian of southeastern China. Asiatyrannus differs from Qianzhousaurus in that it has a proportionally deeper snout, longer premaxilla, deeper maxilla, and deeper dentary, and the cornual process of the lacrimal is inflated without developing a discrete horn. The different skull proportions and body sizes suggest that Asiatyrannus and Qianzhousaurus likely had different feeding strategies and occupied different ecological niches.
Similar content being viewed by others

A new long-snouted marine reptile from the Middle Triassic of China illuminates pachypleurosauroid evolution

A small Cretaceous crocodyliform in a dinosaur nesting ground and the origin of sebecids
A giant tyrannosaur from the Campanian–Maastrichtian of southern North America and the evolution of tyrannosaurid gigantism
Introduction
Tyrannosauroids are the most distinctive, best known, and most intensively studied groups of dinosaurs, and are represented by almost thirty species1,2,3,4,5. The earliest tyrannosaurs appeared in the Middle Jurassic, around 165 million years ago4,6,7,8. They became the apex predators in their respective ecologies during the final 20 million years of the Cretaceous in Asia and western North America9. The colossal body size and deep snout are the characteristics of the ecologically dominant latest Cretaceous tyrannosaurid species9.
The tyrannosaurs in Asia are predominantly found in northern China and Mongolia, with the exception being the long-snouted genus Qianzhousaurus from the Nanxiong Formation (Maastrichtian) in Nankang District of Ganzhou City, southeastern China10. The Nanxiong Formation, which is distributed in Jiangxi and Guangdong provinces, is extremely fossiliferous with a large quantity of dinosaur skeletal remains and dinosaur eggs10,11,12. However, the theropods of the Nanxiong Formation are almost exclusively oviraptorosaurs, with more than seven species of oviraptorosaurs having been reported12,13,14,15,16 with Qianzhousaurus being the only non-oviraptorosaur theropod from the Nanxiong Formation yet described. Currently, Qianzhousaurus is retrieved as belonging to a clade of long-snouted Alioramini within the derived clade of Tyrannosauridae10. Beyond Qianzhousaurus, only some isolated theropod teeth, including possible tyrannosaurid teeth, had been reported from the Nanxiong Formation17. Other than that, several large tyrannosauroid teeth had been reported from the Maastrichtian Dalangshan Formation, Guangdong Province18. The coexistence of the long-snouted Alioramus and the deep-snouted Tarbosaurus are found together at the Tsaagan Khuushu locality in Mongolia, demonstrating that multiple large tyrannosaurids were able to live in sympatry within Asia19.
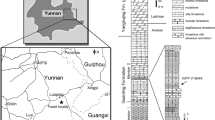
Here, we report the discovery of a new deep-snouted tyrannosaurid from the Upper Cretaceous of Ganzhou City, southeastern China. This new tyrannosaurid specimen was found in a construction site in Shahe Town, Nankang District, Ganzhou City, in September 2017. The specimen was prepared by technicians Chaohe Yu and Anhao Liu in the Zhejiang Museum of Natural History preparation laboratory, Hangzhou, China (ZMNH).
Results
Systematic paleontology
Theropoda Marsh, 1881
Tetanurae Gauthier, 1986
Coelurosauria Huene, 1914
Tyrannosauroidea Osborn, 1905
Tyrannosauridae Osborn, 1905
Tyrannosaurinae Osborn, 1905
Asiatyrannus xui gen. et sp. nov.
Etymology
The generic name is derived from Asia, and the suffix ‘tyrannus’ is derived from the Latin word for ‘king’ or ‘tyrant’, to emphasize that this is the new tyrannosaur collected in the continent of Asia. The specific name honors Dr. Xing Xu (Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences), a distinguished dinosaurologist who contributed greatly to the study of dinosaurs from China, including the research of several tyrannosaurs: Guanlong, Dilong, and Yutyrannus20,21,22. Dr. Xing Xu has also been a great supporter of the paleontological research and science popularization work of the Zhejiang Museum of Natural History.
Holotype
ZMNH M30360, housed at Zhejiang Museum of Natural History, Hangzhou, Zhejiang, China. A nearly complete skull and partial disarticulated postcranial skeleton, including caudal vertebrae; right femur, tibia, and fibula; metatarsals, pedal phalanges, and a partial midshaft of the left tibia, fibula, and metatarsals (Figs. 1, 2, 3, 4, 5, 6 and 7, Table 1).
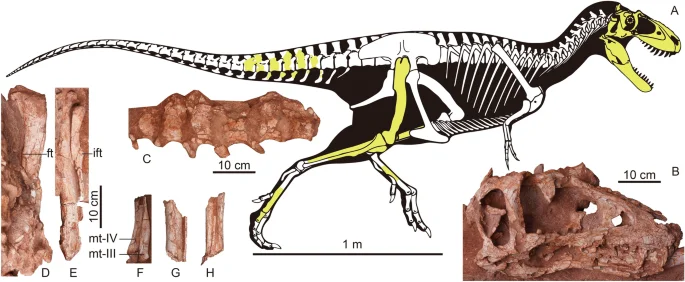
Fossil remains of Asiatyrannus xui (ZMNH M30360). (A) Skeletal outlines showing recovered elements in yellow color. The skeletal reconstruction is the proportional scaling of Lythronax argestes drawn by Scott Hartman from Loewen et al.23; (B) the skull in right lateral view; (C) the caudal vertebrae in left lateral view; (D) the right femur in posterior view; (E) the right tibia and fibula in posterior view; (F) The distal portion of the right metatarsal III in medial view and metatarsal IV in anterior view; (G) the middle shaft of the left tibia and fibula in anterior view; (H) The middle shaft of the left metatarsals in anterior view. ft the fourth trochanter; ift iliofibularis tubercle; mt metatarsal.
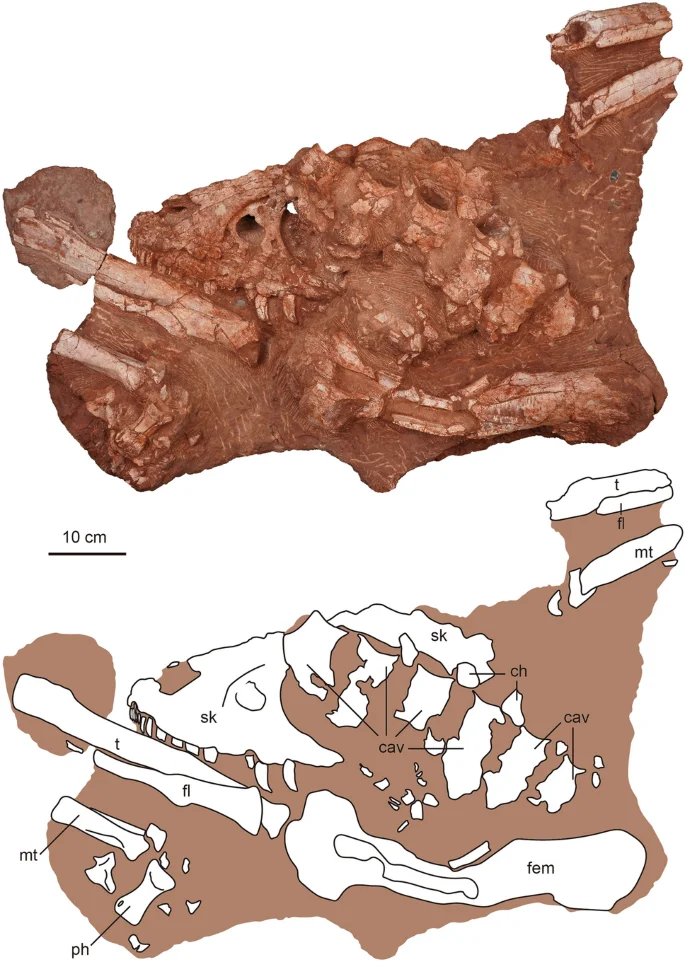
The photography and line drawing of the holotype of Asiatyrannus xui (ZMNH M30360). cav caudal vertebra; ch chevron; fem femur; fl fibula; mt metatarsal; ph phalanx; sk skull; t tibia.
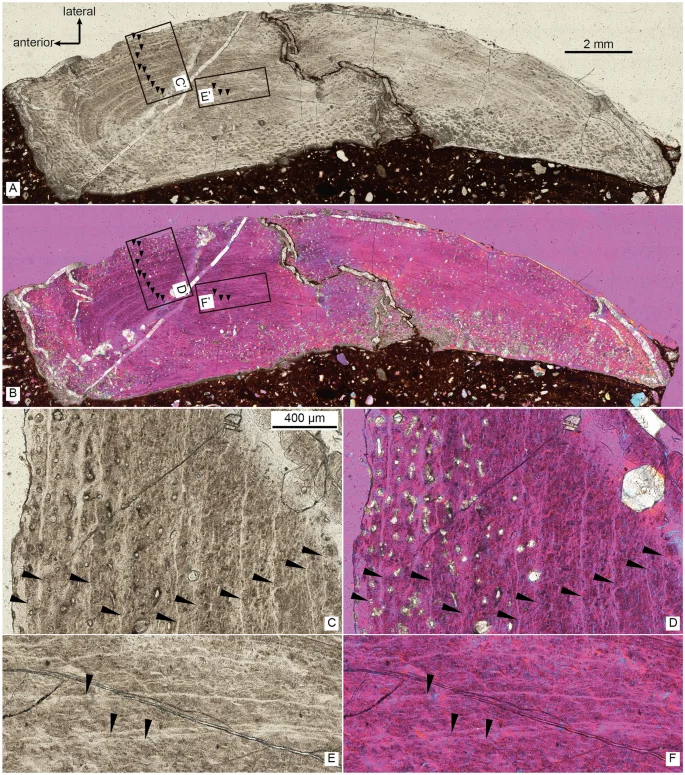
The thin section of the right fibula of Asiatyrannus xui (ZMNH M30360) in normal light (A) and cross-polarized light using a lambda compensator (B), inset boxes C’–F’ indicate the position of the detailed pictures in (C–F); the detail of right fibula in normal light (C, E), and in cross-polarized using a lambda compensator (D, F). Black arrows denote lines of arrested growth (LAGs).
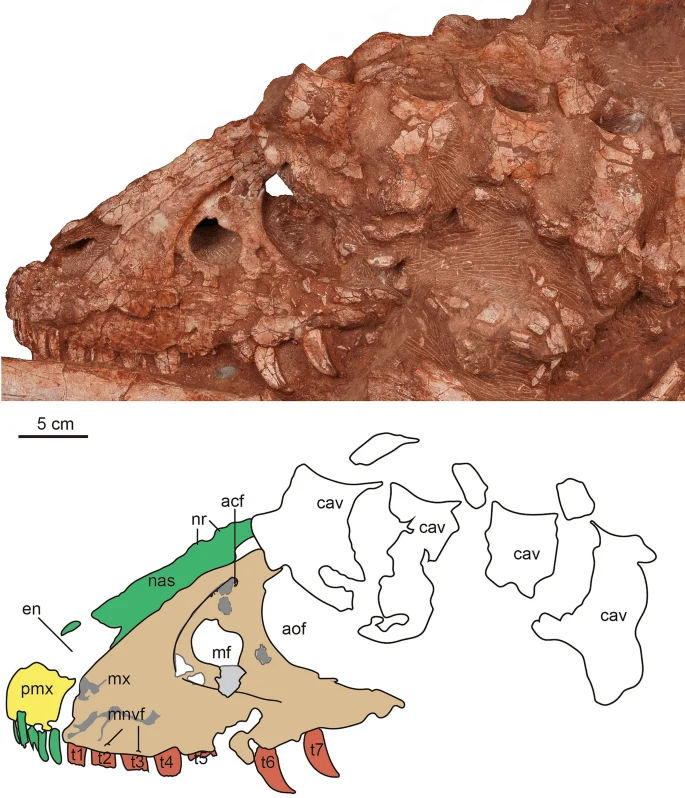
The photograph and line drawing of the skull of Asiatyrannus xui (ZMNH M30360) in left lateral view. acf accessory fossa of maxilla; aof antorbital fenestra; cav caudal vertebra; en external naris; mf maxillary fenestra; mnvf maxillary neurovascular foramina; mx maxilla; nas nasal; nr nasal ridge; pmf promaxillary fenestra; pmx premaxilla; t1–7 maxillary tooth 1–7.
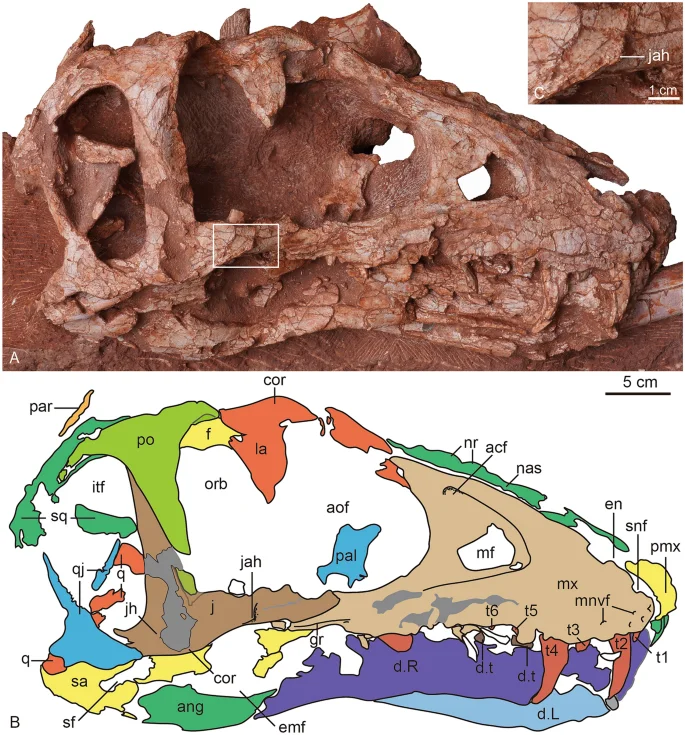
The photograph (A) and line drawing (B) of the skull of Asiatyrannus xui (ZMNH M30360) in right lateral view. The inset box in (A) indicates the position of the detailed jugal accessory horn in (C). acf accessory fossa of maxilla; ang angular; aof antorbital fenestra; cor cornual process; d dentary; d.t dentary tooth; emf external mandibular fenestra; en external naris; f frontal; gr groove; itf infratemporal fenestra; j jugal; jah jugal accessory horn; L left; la lacrimal; mf maxillary fenestra; mnvf maxillary neurovascular foramina; mx maxilla; nas nasal; nr nasal ridge; orb orbit; par parietal; pal palatine; pmx premaxilla; po postorbital; q quadrate; qj quadratojugal; R right; sa surangular; sf surangular foramen; snf subnarial foramen; sq squamosal; t1–6 maxillary tooth 1–6.
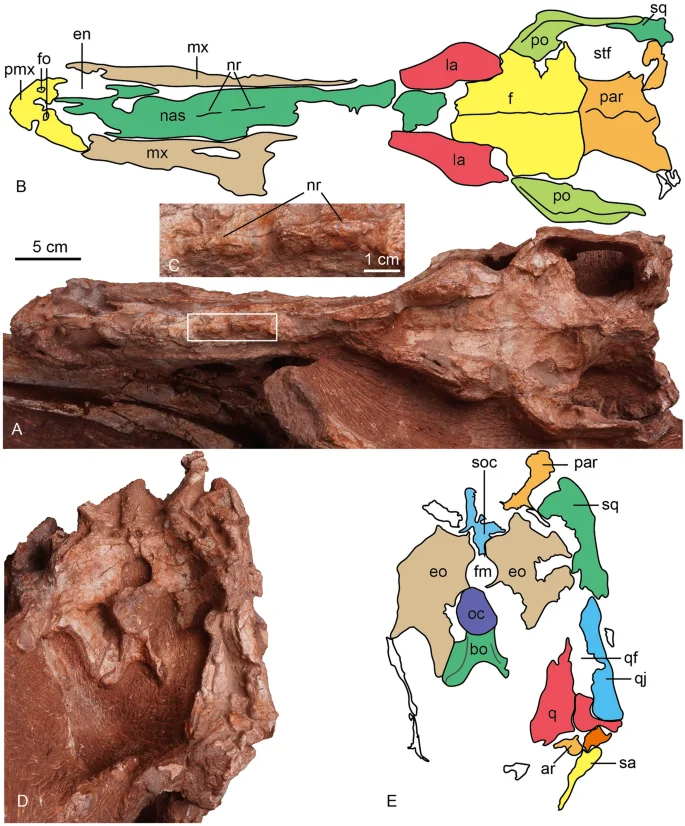
The photograph and line drawing of the skull of Asiatyrannus xui (ZMNH M30360) in dorsal (A, B) and posterior (D, E) views. The inset box in (A) indicates the position of the detailed medium ridges of the nasal in (C). ar articular; bo basioccipital; en external naris; eo exoccipital; fm foramen magnum; fo fossa; la lacrimal; mx maxilla; nr nasal ridge; nas, nasal; oc occipital condyle; par parietal; pmx premaxilla; po postorbital; q quadrate; qf quadrate foramen; qj quadratojugal; sa surangular soc supraoccipital; sq squamosal; stf supratemporal fenestra.
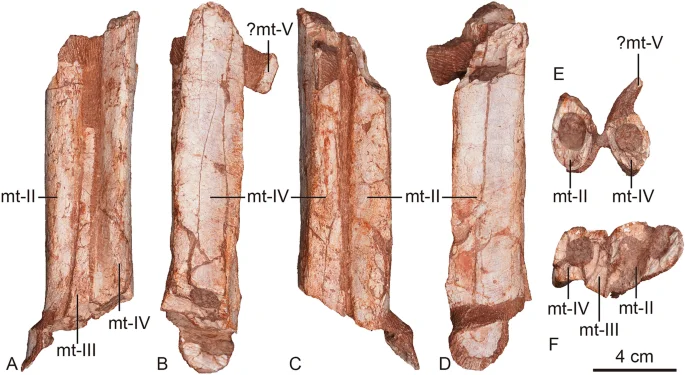
The left metatarsals Asiatyrannus xui (ZMNH M30360) in anterior (A), lateral (B), posterior (C), medial (D), proximal (E), and distal (F) views. mt metatarsal.
Type locality
Nanxiong Formation (Upper Cretaceous, Maastrichtian) of Shahe Town, Nankang District, Ganzhou City, Jiangxi Province, China.
Diagnosis
Asiatyrannus xui is a small to medium-sized tyrannosaurine theropod that differs from other tyrannosauroids in possessing the following autapomorphies: two small, deep fossae located on the lateral surface of the premaxilla just lateral to the anteroventral border of the external naris, a large and sub-rectangular shaped maxillary fenestra, the posterior protuberances of the nasals connect to form two separated medium low ridges, a low ridge-like jugal accessory horn, the ventral margin of the anterior ramus of the jugal curving ventrally anterior to the accessory horn, the lateral surface of the descending process of the postorbital developed the anterodorsally trending fine lineations, the slender, straight, and banded-shaped postorbital bar, with almost straight and parallel anterior and posterior margins in lateral view, and the lateral surangular shelf extends to the posterior end of the surangular.
Ontogenetic age and body size
The periosteal cortex of the right fibula is zonal and exhibits a minimum of 13 lines of arrested growth (LAG) (Fig. 3). The fibular section lacks a medullary cavity similar to the juvenile Tarbosaurus MPC-D 107/724, however, the secondary remodeling is more developed in ZMNH M30360 than MPC-D 107/724. The periosteal bone is incomplete; therefore, the total number of LAGs and the presence of an external fundamental system cannot be determined. Thus, the Asiatyrannus had a terminal age of at least 13 years of life. The outer zones became narrower, indicating a slowdown of growth was occurring, but the animal was still experiencing active growth. The medial half of the fibula had undergone a large degree of secondary remodeling, which obliterated the LAGs (Fig. 3). The narrower outer zone and the highly developed secondary osteons indicate that the holotype of the Asiatyrannus xui is not a full-growth adult individual, but it already passed through the most rapid growth stage. The skull of Asiatyrannus shows several matured cranial morphology: the ventral edge of the maxilla has a distinct convex curvature; the nasals are fully fused with pronounced vaulting and well development of protuberances; the cornual process of lacrimal is strongly inflated similar to the adult Tyrannosaurus and Tarbosaurus2; the dentary is relatively deep. Thus, we interpret the specimen to be a sub-adult that was nearing somatic maturity. The early branching, small-bodied tyrannosauroid Guanlong and Moros reached or neared their adult size at 6–7 years of age20,25. By contrast, the large-bodied, Campano-Maastrichtian tyrannosaurines were undergoing exponential stage growth around 14 years old26. By comparison, the Asiatyrannus holotype is not a full-growth individual and experienced an earlier exponential growth rate when compared with other large-bodied tyrannosaurines from North America26. The holotype’s femoral length is around half the length of individuals with a similar number of LAGs26, thus indicating Asiatyrannus represents a relatively small-bodied tyrannosaurine compared with other large-bodied Campano-Maastrichtian tyrannosaurines. The length of Asiatyrannus skull and femur is 47.5 cm and 44 cm, respectively. The total body length of Asiatyrannus is estimated from 3.5 to 4 m (Fig. 1). The skull is about 8% longer than the femur, while the same ratio in Qianzhousaurus is 29% since the length of the skull and femur is 90 cm and 70 cm, respectively10.
Comparative description
The holotype of the Asiatyrannus xui preserves a nearly complete skull and partial postcranial skeleton. The right side of the skull is well-exposed, but the posterior half of the left side is overlain by the postcranial skeleton. The external texture of most bones is not well-preserved, with the periosteal surface obscured by cracks, breaks, holes, and fractures, resulting from taphonomic processes.
Skull
The posterior half of the left skull is obscured by the caudal vertebrae, and the left mandible is concealed by the hindlimb (Figs. 2, 4), the right side of the skull is well-exposed (Fig. 5). Overall, the skull is well-preserved with all bones articulated with each other, however, some sutures between the bones are difficult to observe due to breakage or poor surface preservation (Figs. 2, 4, 5 and 6). The skull is deeper dorsoventrally than it is transversely wide as in other tyrannosaurids2. The lateral profile of the skull is deeper than alioramins Alioramus27,28, and Qianzhousaurus10, and is similar to other large-bodied tyrannosaurids like Tarbosaurus, Tyrannosaurus, and Gorgosaurus2. The snout and the skull are both relatively narrower than that in large-bodied tyrannosaurids such as Gorgosaurus2, Tarbosaurus29, and Tyrannosaurus29,30. The length of the skull is 47.5 cm from the anterior tip of the premaxilla to the posteriormost point of the occipital condyle, much smaller—but proportionally deeper—than that of Qianzhousaurus, which is 90 cm in length10. In tyrannosaurids, the skull becomes deeper and more robust with increasing maturity or size2,22.
Premaxilla
Both premaxillae are preserved though both the ascending processes are damaged, and the outer surfaces of the premaxillae are broken in areas. The ventral ramus of the premaxilla is taller dorsoventrally than it is long anteroposteriorly, as in other tyrannosauroids2. The length of the ventral surface of the premaxilla is around 6.3–6.9% of the overall basal skull length (Table 1), which is similar to ∼4–10% of other tyrannosauroids, except extremely low 2.2% of Qianzhousaurus10,31. The anterior margin of the premaxilla is oriented almost vertically in lateral view, similar to Tarbosaurus29, and Tyrannosaurus32. This same margin is sloped posterodorsally in ‘Nanotyrannus’1. The premaxilla forms the anterior and ventral edge of the external naris, which is anteroposteriorly long with its long axis sloping anteriorly. Two small, deep fossae are located on the lateral surface of the premaxilla just lateral to the anteroventral border of the external naris (Fig. 6). The fossae, which are present on both premaxillae, are not seen in other tyrannosaurs. Four conical, elongated premaxillary teeth are considerably smaller than the maxillary teeth, as in Xiongguanlong, Timurlengia, and other Late Cretaceous tyrannosauroids2,33. All premaxillary teeth are visible in anterior view, as is characteristic of tyrannosaurids10. Two small foramina are preserved along the ventral margin of the right premaxilla parallel to the tooth row (Fig. 5). The premaxilla is well-separated from the maxilla in lateral view, likely due to the taphonomic distortion, so the location of the subnarial foramen is unclear.
Maxilla
Both maxillae are well-preserved. The ventral margin of the right maxillae is incomplete. The better preserved left maxilla has a distinct and ventrally oriented convex curvature near its midpoint, as seen in Timurlengia and Late Cretaceous large-bodied tyrannosauroids2,32,33, contra to the straight or slightly convex in early-branching tyrannosauroids, such as Guanlong20, Dilong21, Xiongguanlong34, and Suskityrannus33. The curvature of the ventral margin is an ontogenetic feature with this margin having a much weaker convexity in juveniles than in adults, as is seen in Tarbosaurus24,29,35. The curvature of the ventral margin is more strong than in juvenile Tarbosaurus24, but as strong as in large-bodied adult tyrannosauroids, such as Tarbosaurus29, and Tyrannosaurus32. The main body of the maxilla is deep and short as in other large adult tyrannosaurids such as Tyrannosaurus and Tarbosaurus19. Contra with the long and low maxilla in Alioramus remotus28, Alioramus altai19, and Qianzhousaurus10. The promaxillary fenestra is obscured in lateral view, it rotates to face anteriorly as in other tyrannosaurines2. Unlike other tyrannosaurines, the anterior margin of the maxillary fenestra does not contact the anterior rim of the antorbital fossa2. The main body of the maxillae tapers in depth posteriorly below the antorbital fenestra.
Although the outer surface of the maxillae is full of cracks, it is clear the surface does not develop the dorsoventrally trending grooves and ridges present on other tyrannosaurids, as is the case in adult Daspletosaurus (CMN 8506), Tarbosaurus, Tyrannosaurus19, and Zhuchengtyrannus36 specimens. The dorsoventrally trending grooves are present both in juvenile and adult Tarbosaurus24,29. Anteriorly, the maxillae articulate with the premaxilla, and the contact margin is oriented dorsoventrally resulting in a rounded anteroventral corner, as in most tyrannosaurids19. This condition differs from the strong posterodorsal orientation of the premaxillary articulation margin and sharp anteroventral point in Alioramus remotus28, Alioramus altai19, and Qianzhousaurus10. Posteriorly, the main body of the maxilla is articulated with the jugal, with the dorsal half of the maxilla overlaid by the jugal. Some alveolar neurovascular foramina are preserved immediately dorsal and parallel to the tooth row. The antorbital fossa is extensive and extends beyond the antorbital fenestra anteriorly and ventrally, similar to the condition found in Alioramus altai19, and Appalachiosaurus37. However, this condition differs from large adult tyrannosaurids, such as Tyrannosaurus and Tarbosaurus, where there is little lateral exposure of the fossa below the antorbital fenestra19.
The maxillary fenestra is large and sub-rectangular in shape, similar to the adult Tarbosaurus29, in contrast, the fenestra is oval in juvenile Tarbosaurus24. This fenestra is located relatively anteriorly between the anterior edge of the antorbital fossa and fenestra. In contrast, the fenestra is located more centrally along the maxilla of Alioramus altai, Appalachiosaurus, Albertosaurus, Bistahieversor, Gorgosaurus, and juvenile tyrannosaurines19. The margin of the maxillary fenestra is widely separated from that of the external antorbital fenestra, as in small-bodied tyrannosauroids such as Raptorex38, and juvenile specimens of tyrannosaurids, like Tarbosaurus24. Although the skull is smaller, the fenestra is relatively larger than in Appalachiosaurus37, juvenile and adult Gorgosaurus39,40, and ‘Nanotyrannus’1. The maxillary tooth row is not completely preserved. The first tooth of the right maxilla is small and incisiform, similar to the premaxillary teeth (Fig. 5), but the first tooth of the left maxilla seems as large as the second one (Fig. 4).
Nasal
The left and right nasals are fused medially into a single vaulted element, a characteristic feature of tyrannosauroids19,41. Anteriorly each nasal divides into two processes: the subnarial process and the premaxillary process, as in other tyrannosaurs19. Anteriorly, the premaxillary processes are separated on the midline. The nasal is excluded from the antorbital fossa by the maxilla and lacrimal, in contrast, the nasal forms a part of this margin in adult Tarbosaurus24,29. In dorsal view, the subnarial processes are lateral to the premaxillary processes. This condition is also seen in Gorgosaurus, Albertosaurus, some specimens of Tyrannosaurus (BHI 3033), and Alioramus altai19. In contrast, the subnarial process is nearly beneath the premaxillary process in Daspletosaurus (CMN 8506) and other specimens of Tyrannosaurus (CM 79057)19. The nasals don’t show any separation in the anterior end. However, they are separated at the anterior and posterior ends in juvenile and adult Tarbosaurus24. The dorsal margin is straight in lateral view, in contrast, the anterior half of the dorsal margin is vaulting in lateral view in adult Tarbosaurus29.
The nasal has pronounced vaulting observed in large tyrannosaurids42. The dorsal external surface of the nasal is rugose with prominent developed nasal protuberances, as is the case in most other tyrannosaurids2,32. The protuberances are homologous to the rugosities seen in all Late Cretaceous tyrannosauroids, Alioramus remotus28, Appalachiosaurus37, Yutyrannus22,34, except Xiongguanlong34. The rugose region is developed in all preserved nasals and extends around the middle of the antorbital fenestra. The posterior protuberances are connected to form two successive separated low medium ridges (Figs. 4, 5 and 6: nr). The development of the nasal protuberances is an ontogenetic feature and the nasal becomes more pronounced in larger specimens2. The dorsal surface of the nasal is smooth in Raptorex38, and the juvenile Tarbosaurus24. The pronounced protuberances on the fused nasal of Asiatyrannus are similar to other adult tyrannosaurids, such as Tarbosaurus29, Tyrannosaurus32, and Qianzhousaurus10.
Lacrimal
Both both lacrimals are preserved, but the caudal vertebrae obscure the left and the right is incomplete (Figs. 5, 6). The bone is L-shaped in lateral view, with a short posterior process that is similar to that of Tyrannosaurus1. The angle between the anterior and ventral rami is acute, similar to Daspletosaurus, Tarbosaurus, Tyrannosaurus, and Alioramus altai19. The cornual process is inflated without a discrete hornlet similar to the Tyrannosaurus–Tarbosaurus clade2. The cornual process is inflated similar to that of Tyrannosaurus32, and is more inflated than in Appalachiosaurus37, and the adult specimen of Tarbosaurus19,29. However, the surface of the cornual process is smoother than that of Tarbosaurus29. The inflation is not developed in Raptorex38, or less pronounced in the juvenile specimen of Tarbosaurus24. The cornual process has a triangular horn in Yutyrannus22, albertosaurines2,40, Daspletosaurus2, Alioramus altai19, and Qianzhousaurus31.
Jugal
The lineation between the postorbital and jugal is a straight, obliquely angled line. This contact divides the postorbital bar diagonally in an anteroventral to posterodorsal direction. However, the line is distinctly bent in Daspletosaurus, Albertosaurus2, and Tarbosaurus29. The ascending process terminates at the anterodorsal rim of the infratemporal fenestra but does not contact the squamosal. The ascending process meets the squamosal and excludes the postorbital from the infratemporal fenestra anteriorly in the Tyrannosaurus32. The lateral surface of the ascending process is smooth, as in Albertosaurus, Bistahieversor, Gorgosaurus, Teratophoneus, juvenile tyrannosaurines, and Alioramus altai19. By contrast, this lateral surface is excavated by a broad concavity in large subadult and adult tyrannosaurines (Daspletosaurus, Tarbosaurus, Tyrannosaurus)19. The entire postorbital contact zone is demarcated posteriorly by a pronounced ridge, as in Alioramus altai19. The ridge is especially prominent in the dorsal half of the jugal.
Posterior to the ascending process, the jugal forms the ventral floor of the infratemporal fenestra. The posterior ascending process of the jugal has an acute dorsal tip and is overlapped by the postorbital anteriorly, as in Tyrannosaurus32. The jugal horn appears to be developed, but the broken surface of the horn makes the condition uncertain. The jugal horn is only well developed in Alioramus altai19, and ridge-like in Qianzhousaurus31. Similar to Alioramus altai19, the lateral surface of the jugal’s external surface develops anteroposterior raised lineations. Along the ventral margin of the anterior ramus and ventral to the orbit, there is a ridge-like jugal accessory horn in the ventrolateral portion of the anterior process (Fig. 5). The accessory horn is not present in other tyrannosauroids including Raptorex38, the juvenile and adult of Tarbosaurus24,29. Anteriorly, the ventral margin of the anterior ramus has a ventral curvature anterior to the accessory horn. However, the anterior ventral margin is straight or curves slightly dorsally in other tyrannosauroids2,30, including juvenile and adult Tarbosaurus24,29.
Frontal
Both frontals are well-preserved (Figs. 5, 6). The frontals are sutured medially as in other tyrannosaurids2. In dorsal view, the contact with the postorbital is V-shaped, with a concavity located on the frontal, as in Tyrannosaurus32. However, the border between the prefrontal and the prefrontal is unclear. The posterior portion of the frontals slopes ventrally into the supratemporal space from the anterior and medial supratemporal borders with a low sagittal crest extending posteriorly to parietals. The supratemporal fossae in the frontals are short, as in ’Nanotyrannus’, whereas they are elongated in Tyrannosaurus1. There is only a short contribution of frontal to the sagittal crest, like in ‘Nanotyrannus’. In contrast, there is a long frontal contribution to the sagittal crest in Tyrannosaurus1. The frontal has a small, notch-like orbital rim, as in later-diverging taxa33. In contrast, the frontal makes a wide contribution to the orbital rim in Guanlong, Dilong, and Suskityrannus33.
Parietal
As in other tyrannosaurids, the parietals possess a fused sagittal crest2,32. The nuchal crest is broken and not present along the parietals. The frontoparietal suture is distinct and transversely directed. The parietal slopes broadly laterally within the supratemporal space, forming the medial and medioposterior walls of the fenestrae.
Postorbital
The right postorbital is well-preserved and exposed, while the lateral surface of the left postorbital is obscured by caudal vertebrae. Although the anterior portion of the process is broken, the cornual process is swollen and is present as a rugose convex boss that is located posterodorsally to the orbit, as in mature specimens of all other tyrannosaurids31. This process is absent in basal tyrannosauroids, but present as well-developed, rugose bulges in tyrannosaurids19. The descending process is tongue-shaped and gracile, as in Alioramus altai19 and ‘Nanotyrannus’1. The descending process is straight and gracile, without an anteriorly expanded suborbital process. The descending process is anteroposteriorly broad and constricts the orbit in large individuals of Yutyrannus22, Tarbosaurus29, Tyrannosaurus1, and Albertosaurus43. The anterior expansion is incipient in the juvenile specimen of Tarbosaurus24, however, it is more conspicuous in the juvenile specimen of Tarbosaurus than in Asiatyrannus24. Although the anterior expansion is incipient in Gorgosaurus, the descending process curved anteriorly in juvenile and adult individuals39,40, in contrast, the anterior edge of Asiatyrannus is almost straight. The descending process reaches the floor of the orbit ventrally, however, the process terminated higher in juvenile and adult Tarbosaurus24,29. The lateral surface of the process has anterodorsally trending fine lineations. In contrast, the lineations are horizontally directed and dorsally convex in Alioramus altai19, Albertosaurus39, Qianzhousaurus31, Tarbosaurus24, and ‘Nanotyrannus’1. The postorbital bar composed of the jugal and postorbital is slender, straight, and banded-shaped, with almost straight and parallel anterior and posterior margins in lateral view. Generally, the postorbital bar is broader and the anterior margin is convex anteriorly due to the development of the anterior expansion of the postorbital. In other tyrannosauroids without anterior expansion development, the anterior margin is concave anteriorly, such as Raptorex38, Daspletosaurus30, ‘Nanotyrannus’1, and juvenile Tarbosaurus24.
Squamosal
The squamosal is broadly convex and slopes posteroventrally in lateral view. The anterolateral process is forked, with dorsal and ventral projections. The lateral surface is overlapped by the posterior ramus of the postorbital. In lateral view, the dorsal projection of the squamosal is exposed and is wider than the ventral projection. The descending process forms the derived squamosal-quadratojugal flange that bisects the infratemporal fenestra, which may be a tyrannosaurid synapomorphy2,32,41. The squamosal component contacts with the postorbital bar anteriorly and bisects the infratemporal fenestra completely, as in Tyrannosaurus32, but not in juvenile and adult Gorgosaurus39,40, juvenile and adult Tarbosaurus24,29, Alioramus altai19, and Qianzhousaurus31. The anterior tip of the flange is rugose and bulbous. The long axis of the ventral process projects roughly anteroposteriorly, as in other tyrannosaurids19. Posteroventrally, a portion of the squamosal is broken, however, the preserved portion of the posterior end directs posteroventrally. In the posterodorsal region of the lateral margin, there is a prominent thin ridge that extends along the dorsal edge. The crest directs dorsally and marks the highest point of the squamosal, as in Tyrannosaurus32. However, the ridge is located along the lateral surface of the squamosal rather than its dorsal edge as in Tarbosaurus29 and Alioramus altai19.
Quadratojugal
The anterior and posterior margins of the quadratojugal’s ascending processes are both concave, making the quadratojugal hourglass-shaped in lateral view. This condition is similar to that observed in other tyrannosaurids19. The posterior process is short and directs ventroposteriorly. In lateral view, the ventral edge of the quadratojugal is strongly concave.
Quadrate
The quadrate forms the mandibular condyle, which is transversely expanded. The condyle is extended posterior to the posteriormost extent of the paroccipital processes, contra to other tyrannosaurids2.
The occipital elements
The occipital condyle is incomplete. The sutural separation of the exoccipitals and basioccipital is only visible ventrally, lateral to the basal tubera. The paroccipital processes are broad. The basal tubera is deep and bilaterally narrow as in Daspletosaurus sp. (TMP 94.143.1)39, however, the basal tubera is broad in Tyrannosaurus32. The basal tubera is separated along its ventral margins by a concave notch that is deep and more than half of the depth of the tubera—similar to the derived tyrannosauroids44, Alioramus remotus28, and Tarbosaurus29. In contrast, this notch is shallow in Tyrannosaurus32, and Qianzhousaurus31, or narrow in ‘Nanotyrannus’1.
Mandible
The mandible is preserved in a closed position with the cranium, thus making the dorsal portion of the mandible obscured by the cranium. The mandible is robust and deep as in those of adult tyrannosaurids. The dentary is more robust and deeper dorsoventrally than those of alioramins, such as Alioramus27, and Qianzhousaurus10.
Both dentaries are preserved with most of the lateral right dentary and ventral portion of the medial left dentary exposed. The anterior edge of both dentaries is more vertically directed than those of alioramins, such as Alioramus27, and Qianzhousaurus10. The ventral edge of the dentary has only slight concave curvature, such as Zhuchengtyrannus36.
The surangular is badly damaged along the dentary and posterior margins of the external mandibular foramen. Dorsal to the surangular foramen, the lateral surangular shelf is prominent and overhangs its lateral surface, as in Tyrannosaurus32. The shelf extended to the posterior end. In contrast, the edge of the surangular ends posteriorly above the posterodorsal corner of the surangular foramen in ‘Bagaraatan’45, Tyrannosaurus32, and Alioramus altai19.
The angular is also badly damaged. The anterior dorsal edge, which borders the ventral of the external mandibular fenestra, appears complete and has a convex lateral surface19. The ventral edge of the angular forms the ventral edge of the mandible and is convex in lateral view. The ventral border of the angular is broadly convex, and there are no prominent rugosities as reported in Tyrannosaurus19,32.
Postcranial skeleton
Femur
The right hindlimb is well-preserved, with the femur, tibia, fibula, and partial pes preserved (Fig. 1). Only a partial tibia, fibula, and metatarsals from the left hindlimb were recovered (Figs. 2, 7). As in most theropods, including tyrannosauroids, the femur has a posteriorly oriented concave curve in lateral view (Fig. 2)2,25. The head of the femur projects mostly medially but also slightly proximally (Fig. 1). The proximal end of the right femoral head and the greater trochanter are partially missing due to the eroded nature of the proximal femur. Based on the preserved femur, the head is not distinctly elevated relative to the greater trochanter. Due to the incompleteness of the lesser trochanter, the relative height of the lesser and greater trochanters is indeterminate. They were separated by a deep, narrow cleft. The medullary cavity is expansive and cortical bone is relatively thin as in other tyrannosauroids25. In proximal view, the femoral head keeps a relatively constant width as it extends medially, similar to Gorgosaurus and Tarbosaurus19. A well-developed sulcus that would have housed ligaments is present along the midline of the posterior surface of the femoral head, as is observed in Alioramus altai19. The subtriangular fourth trochanter is powerfully developed along the posteromedial shaft, as in other tyrannosauroids2,25.
Tibia
Both tibiae are incomplete, with the right tibia being the most complete with only the distal articular condyles missing (Figs. 1, 2). The preserved length of the right tibia is shorter than the length of the right femur. Extensive hollowing is found throughout the tibia with the wall of the tibia being thin and the marrow cavity being relatively large.
Fibula
The fibular shaft is tightly appressed to the tibia as in Bistahieversor, and Moros25. The proximal half of the right fibula and the midshaft of the left fibula are preserved, with both articulating with the corresponding tibia (Figs. 1, 2). The iliofibularis tubercle is pronounced on the margin where the anterior and lateral surfaces meet, This iliofibularis tubercle, is large, prominent, rugose, and elongated proximodistally as in most tyrannosauroids like Dryptosaurus46. The fibular shaft is most constricted distal to the iliofiburlaris tubercle in lateral view.
Metatarsus and pes
The left and right metatarsals are preserved in articulation but are incomplete, with the mid-shaft section of the left metatarsal II–IV and the distal portion of right metatarsal III and IV being preserved (Figs. 1, 7). As in other tyrannosauroids, the metatarsus is long and slender2. The metatarsus is arctometatarsalian, with the metatarsal III pinched between metatarsals II and IV proximally, as in all tyrannosaurids2,19, and some non-tyrannosaurids such as Appalachiosaurus37, Bistahieversor, Dryptosaurus46, and Raptorex38. In contrast, a more normal theropod metatarsus, in which metatarsal III is large and un-pinched, is seen in the early branching tyrannosauroids, Kileskus7, Dilong21, Eotyrannus47, Guanlong20, and Yutyrannus22. The pinched proximal shaft of the metatarsal III is visible in anterior view. The whole preserved portion of metatarsal III is unobservable in posterior view, with metatarsal III and IV almost contacting each other. This condition where metatarsal III is proximally constricted is observed in tyrannosaurids and some non-tyrannosaurid tyrannosauroids. However, the proximal end of metatarsal III does not reach the proximal articulate surface with the tarsus, most likely due to poor preservation. The right metatarsal III is exposed in medial view, though its proximal portion missing. The preserved proximal portion is narrow in medial view, and it flares smoothly distally where it reaches its great width around the midpoint of the preserved shaft. The left surface shaft is inflated. The shaft has a constricted neck just proximal to the condyle. The medial ligament pit is large, deep, circular, and well-developed on the medial side of the condylar region.
Some nondescript pedal phalanges are preserved disarticulated and located beside the right metatarsus, with one exposed in dorsal view and two exposed in lateral view. They are considered to be from the right pes since they are non-ungual phalanges. In the dorsal exposed phalanx, it is constricted in the middle, with expanded proximal and distal articular regions.
Discussion
The phylogenetic analysis resulted in the recovery of 8 MPTs, (tree length = 861; consistency index = 0.521; retention index = 0.793), the strict consensus of which places Asiatyrannus in the most derived tyrannosauroid clade–Tyrannosaurinae (Fig. 8).
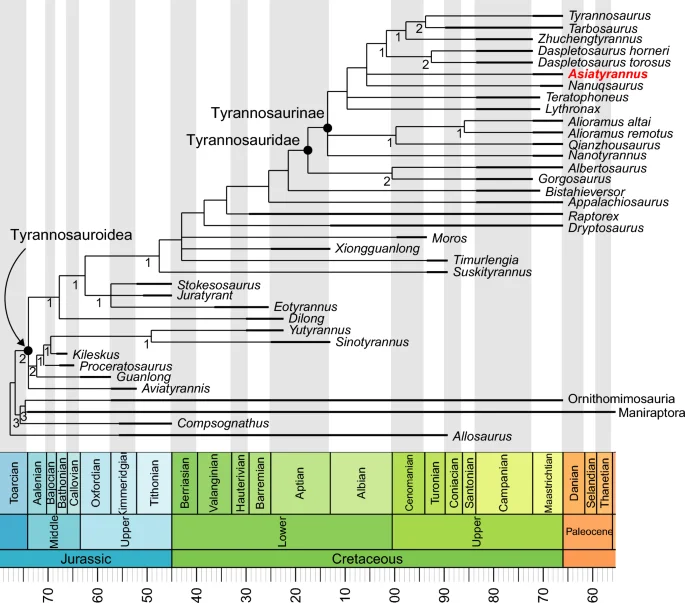
The strict consensus tree produced by phylogenetic analysis. Values beneath nodes indicate Bremer support.
Asiatyrannus is a tyrannosaurid theropod based on its possessing the following features: four incisiform premaxillary teeth, fused nasals, infratemporal fenestra constricted by the squamosal-quadratojugal flange, and the arctometatarsalian foot2. Both Asiatyrannus and Qianzhousaurus are discovered in the Nanxiong Formation of Nankang District, Ganzhou City10. Asiatyrannus differs from Qianzhousaurus with a proportionally deeper snout, longer premaxilla, deeper maxilla, and deeper dentary, and the cornual process of the lacrimal is inflated without a discrete horn10. Generally, the skulls of tyrannosauroids become deeper as their size increases, however, the Asiatyrannus skull (47.5 cm) is only around half in length as Qianzhousaurus (90 cm) but is proportionally deeper. Asiatyrannus is the first definitive deep-snout tyrannosaurid dinosaur from southern China and is the southernmost occurrence of tyrannosaurids in Asia.
The phylogenetic analysis places Asiatyrannus xui as a derived tyrannosaurid deeply nested in Tyrannosaurinae, positioned apart from the long-snouted alioramins and other early branching tyrannosaurines, and closer to the largest deep-snouted Tarbosaurus, Tyrannosaurus, and Zhuchengtyrannus (Fig. 8). In the latest Cretaceous, tyrannosaurids became restricted to Asia and North America, and they developed into large-bodied, bone-crunching apex predators—as exemplified by Tarbosaurus, Tyrannosaurus, and Zhuchengtyrannus4,33,36. Among tyrannosaurids, ‘Nanotyrannus’ (skull length 57.2 cm) and Nanuqsaurus (estimated skull length 60–70 cm in mature individuals), both larger than Asiatyrannus, might also represent the small to medium-bodied tyrannosaurids48,49. However, the taxonomic status of ‘Nanotyrannus’ is highly controversial, with the interpretation of ‘Nanotyrannus’ as a juvenile Tyrannosaurus being widely accepted1,30,48,50. Thus Nanuqsaurus might be the small-bodied tyrannosaurid that inhabited in high-latitude of North America49. In seven of our eight MPTs, Asiatyrannus and Nanuqsaurus were recovered as sister taxa, however, only one character of the small body size (char 1) grouped them together. New material from the Prince Creek Formation suggests Nanuqsaurus has an adult body size more closely comparable to other North American tyrannosaurid taxa, such as Albertosaurus51. Thus, Asiatyrannus currently represents the only definitive small to medium-bodied tyrannosaurine.
More than one tyrannosaurids were discovered in several well-sampled assemblages, such as the Dinosaur Park Formation, the Judith River Formation, and the Nemegt Formation1. Asiatyrannus is part of the diversification of deep-snouted tyrannosaurids and emphasizes the higher species richness of theropods, especially tyrannosaurids, in southeastern China. The discovery reveals the coexistence of the long-snouted and deep-snouted tyrannosaurid in southeastern China as in the Late Cretaceous of central Asia27. However, the body size of the deep-snouted and long-snouted tyrannosaurids is the opposite in southeastern Asia compared to central Asia. Asiatyrannus is a small to medium-sized tyrannosaurine, with an estimated total body length of approximately 3.5–4 m. The histological study demonstrated that the individual is not a fully growth adult, however, Asiatyrannus is less than half the size of the other large-bodied tyrannosaurines in similar growth stages. Also, Asiatyrannus is only half the size of the coeval long-snouted Qianzhousaurus10. Asiatyrannus and Qianzhousaurus have different skull proportions and body sizes, suggesting they may occupy different ecological niches. In the Campano-Maastrichtian of eastern/central Asia and Laramidia, the large carnivore guilds are monopolized by tyrannosaurids, with adult medium-sized predators rare or absent52. Holtz52 interpreted the “missing middle-sized” niches in the theropod guilds of Late Cretaceous Laramidia and Asia may have been assimilated by juvenile and subadults of tyrannosaurid species. In southeastern China, the Qianzhousaurus undoubtedly occupied the apex predator10, but Asiatyrannus might represent the small to medium-sized theropod niche between the large-bodied Qianzhousaurus and the diversified small-bodied oviraptorosaurs.
Methods
Histological sectioning
To assess the terminal age, growth rate, and ontogenetic stage of the holotype of Asiatyrannus xui (ZMNH M30360), we processed a histological thin section from the mid-diaphyseal of the right fibula. We prepared the histological thin section in the Key Laboratory of Vertebrate Evolution and Human Origin of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences (IVPP). The section methodology followed the procedure described by Zhao et al.53. The samples were impregnated with resin and placed in a vacuum chamber to reduce the formation of air bubbles for approximately 12 h and were then embedded in Technovit 7200 VLC light curing 1-component-resin, and cut transversely at the mid-diaphysis of the limb bones. Sectioned surfaces were smoothed with grinding powder and afterward glued onto a glass slide with Araldite and ground to a thickness of 60 to 100 μm. Finally, a glass cover slip for sample protection. The sections were examined in normal light and cross-polarized light and imaged using the Zeiss Axio Imager 2 MAT Upright Microscope with Zen pro 3.7 software in IVPP.
Phylogenetic analysis
To determine the systematic position of Asiatyrannus xui, we used the dataset that focused on the tyrannosauroid evolution54, which is based on the dataset of Brusatte and Carr3. We added Moros25, Suskityrannus33, and Asiatyrannus to the dataset for analysis. Although the status of ‘Nanotyrannus’ currently remains unresolved, we added it to our analysis. and we coded ‘Nanotyrannus’ following Longrich and Saitta1. The revised dataset includes 37 taxa and 386 characters. We assembled the dataset in Mesquite V3.8155, and analyzed the dataset in TNT version 1.6 under maximum parsimony56, with 49 characters (1, 6, 7, 13, 14, 15, 18, 24, 34, 37, 42, 50, 53, 73, 82, 83, 98, 103, 104, 110, 116, 126, 132, 133, 148, 155, 159, 160, 161, 209, 240, 241, 244, 251, 257, 259, 264, 283, 290, 305, 307, 323, 345, 353, 359, 361, 362, 366, 383) ordered following Carr et al.54. We first analyzed the data matrix under the ‘New Technology’ search options, using sectorial search, ratchet, tree drift, and tree fuse options with default parameters. The stored trees were then analyzed in ‘Traditional Analysis’ under the tree bisection reconnection branch (TBR) swapping algorithm.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank. The LSIDs are D0B4FE1C-F558-4A0D-B524-20191D890A9B for this publication, 0248BA77-3106-4E7F-83A4-C67EA6777A54 for the new genus Asiatyrannus, and BE376950-B55A-40FE-8A1D-5E5081803422 for the new species Asiatyrannus xui.
Data availability
Data is provided within the manuscript or supplementary information file.
References
-
Longrich, N. R. & Saitta, E. T. Taxonomic status of Nanotyrannus lancensis (Dinosauria: Tyrannosauroidea)—A distinct taxon of small-bodied tyrannosaur. Fossil Stud. 2, 1–65. https://doi.org/10.3390/fossils2010001 (2024).
Google Scholar
-
Holtz, T. R. J. in The Dinosauria (second edition) (eds D. B. Weishampel, P. Dodson, & H Osmólska) 111–136 (University of California Press, 2004).
-
Brusatte, S. L. & Carr, T. D. The phylogeny and evolutionary history of tyrannosauroid dinosaurs. Sci. Rep. 6, 20252. https://doi.org/10.1038/srep20252 (2016).
Google Scholar
-
Brusatte, S. L. et al. Tyrannosaur paleobiology: New research on ancient exemplar organisms. Science 329, 1481–1485. https://doi.org/10.1126/science.1193304 (2010).
Google Scholar
-
Holtz, T. R. J. The phylogenetic position of the Tyrannosauridae: Implications for theropod systematics. J. Paleontl. 68, 1100–1117. https://doi.org/10.1017/S0022336000026706 (1994).
Google Scholar
-
Rauhut, O. W. M., Milner, A. C. & Moore-Fay, S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zool. J. Linnean Soc. 158, 155–195. https://doi.org/10.1111/j.1096-3642.2009.00591.x (2010).
Google Scholar
-
Averianov, A. O., Krasnolutskii, S. A. & Ivantsov, S. V. A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia. Proc. Zool. Inst. 314, 42–57. https://doi.org/10.31610/trudyzin/2010.314.1.42 (2010).
Google Scholar
-
Averianov, A. O. et al. New data on the tyrannosauroid dinosaur Kileskus from the Middle Jurassic of Siberia, Russia. Hist. Biol. 33, 897–903. https://doi.org/10.1080/08912963.2019.1666839 (2021).
Google Scholar
-
Brusatte, S. L., Averianov, A., Sues, H.-D., Muir, A. & Butler, I. B. New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. Proc. Natl. Acad. Sci. 113, 3447–3452. https://doi.org/10.1073/pnas.1600140113 (2016).
Google Scholar
-
Lü, J. et al. A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nat. Commun. 5, 3788. https://doi.org/10.1038/ncomms4788 (2014).
Google Scholar
-
Wang, S., Zhang, S., Sullivan, C. & Xing, X. Elongatoolithid eggs containing oviraptorid (Theropoda, Oviraptorosauria) embryos from the Upper Cretaceous of Southern China. BMC Evolut. Biol. 16, 67–88. https://doi.org/10.1186/s12862-016-0633-0 (2016).
Google Scholar
-
Wang, S., Sun, C., Sullivan, C. & Xu, X. A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of southern China. Zootaxa 3640, 242–251. https://doi.org/10.11646/zootaxa.3640.2.7 (2013).
Google Scholar
-
Xu, X. & Han, F. A new oviraptorid dinosaur (Theropoda: Oviraptorosauria) from the Upper Cretaceous of China. Vertebr. Palasiat. 48, 11–18 (2010).
-
Lü, J., Yi, L., Zhong, H. & Wei, X. A New Oviraptorosaur (Dinosauria: Oviraptorosauria) from the Late Cretaceous of Southern China and Its Paleoecological Implications. Plos One 8, e80557. https://doi.org/10.1371/journal.pone.0080557 (2013).
Google Scholar
-
Lü, J. C. Oviraptorid Dinosaurs from Southern China (Gelogical Publishing House, 2005).
-
Lü, J. et al. High diversity of the Ganzhou Oviraptorid Fauna increased by a new “cassowary-like” crested species. Sci. Rep. 7, 6393. https://doi.org/10.1038/s41598-017-05016-6 (2017).
Google Scholar
-
Mo, J. & Xu, X. Large theropod teeth from the Upper Cretaceous of Jiangxi, southern China. Vertebr. Palasiat. 53, 63–72 (2015).
-
Xing, L. et al. Large theropod teeth from the Upper Cretaceous of Guangdong Province, Southern China. Cretac. Res. 161, 105914. https://doi.org/10.1016/j.cretres.2024.105914 (2024).
Google Scholar
-
Brusatte, S. L., Carr, T. D. & Norell, M. A. The Osteology of Alioramus, a gracile and long-snouted tyrannosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Mongolia. Bull. Am. Museum Nat. Hist. 366, 1–197. https://doi.org/10.1206/770.1 (2012).
Google Scholar
-
Xu, X. et al. A basal tyrannosauroid dinosaur from the Late Jurassic of China. Nature 439, 715–718. https://doi.org/10.1038/nature04511 (2006).
Google Scholar
-
Xu, X. et al. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431, 680–684. https://doi.org/10.1038/nature02855 (2004).
Google Scholar
-
Xu, X. et al. A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature 484, 92–95. https://doi.org/10.1038/nature10906 (2012).
Google Scholar
-
Loewen, M. A., Irmis, R. B., Sertich, J. J. W., Currie, P. J. & Sampson, S. D. Tyrant dinosaur evolution tracks the rise and fall of Late Cretaceous oceans. PLOS ONE 8, 1–14. https://doi.org/10.1371/journal.pone.0079420 (2013).
Google Scholar
-
Tsuihiji, T. et al. Cranial osteology of a juvenile specimen of Tarbosaurus bataar (Theropoda, Tyrannosauridae) from the Nemegt Formation (Upper Cretaceous) of Bugin Tsav, Mongolia. J. Vertebr. Paleontol. 31, 497–517. https://doi.org/10.1080/02724634.2011.557116 (2011).
Google Scholar
-
Zanno, L. E. et al. Diminutive fleet-footed tyrannosauroid narrows the 70-million-year gap in the North American fossil record. Commun. Biol. 2, 64. https://doi.org/10.1038/s42003-019-0308-7 (2019).
Google Scholar
-
Erickson, G. M. et al. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430, 772–775. https://doi.org/10.1038/nature16487 (2004).
Google Scholar
-
Brusatte, S. L., Carr, T. D., Erickson, G. M., Bever, G. S. & Norell, M. A. A long-snouted, multihorned tyrannosaurid from the Late Cretaceous of Mongolia. Proc. Natl. Acad. Sci. 106, 17261–17266. https://doi.org/10.1073/pnas.0906911106 (2009).
Google Scholar
-
Kurzanov, S. M. A new Late Cretaceous carnosaur from Nogon-Tsav Mongolia. Sovmestnaâ Sovetsko-Mongolskaâ Paleontologiceskaâ Ekspeditciâ, Trudy 3, 93–104 (1976).
-
Hurum, J. H. & Sabath, K. Giant theropod dinosaurs from Asia and North America: skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. Acta Palaeontol. Pol. 48, 161–190 (2003).
-
Carr, T. D. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). J. Vertebr. Paleontol. 19, 497–520. https://doi.org/10.1080/02724634.1999.10011161 (1999).
Google Scholar
-
Foster, W. et al. The cranial anatomy of the long-snouted tyrannosaurid dinosaur Qianzhousaurus sinensis from the Upper Cretaceous of China. J. Vertebr. Paleontol. 41, e1999251. https://doi.org/10.1080/02724634.2021.1999251 (2022).
Google Scholar
-
Brochu, C. A. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. J. Vertebr. Paleontol. Mem. 7, 1–138. https://doi.org/10.1080/02724634.2003.10010947 (2003).
Google Scholar
-
Nesbitt, S. J. et al. A mid-Cretaceous tyrannosauroid and the origin of North American end-Cretaceous dinosaur assemblages. Nat. Ecol. Evolut. 3, 892–899. https://doi.org/10.1038/s41559-019-0888-0 (2019).
Google Scholar
-
Li, D., Norell, M. A., Gao, K. Q., Smith, N. D. & Makovicky, P. J. A longirostrine tyrannosauroid from the Early Cretaceous of China. Proc. R. Soc. B Biol. Sci. 277, 183. https://doi.org/10.1098/rspb.2009.0249 (2010).
Google Scholar
-
Currie, P. J. & Dong, Z. New information on Shanshanosaurus huoyanshanensis, a juvenile tyrannosaurid (Theropoda, Dinosauria) from the Late Cretaceous of China. Can. J. Earth Sci. 38, 1729–1737. https://doi.org/10.1139/e01-042 (2001).
Google Scholar
-
Hone, D. W. E. et al. A new, large tyrannosaurine theropod from the Upper Cretaceous of China. Cretac. Res. 32, 495–503. https://doi.org/10.1016/j.cretres.2011.03.005 (2011).
Google Scholar
-
Carr, T. D., Williamson, T. E. & Schwimmer, D. R. A new genus and species of tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama. J. Vertebr. Paleontol. 25, 119–143. https://doi.org/10.1671/0272-4634(2005)025[0119:angaso]2.0.co;2 (2005).
Google Scholar
-
Sereno, P. C. et al. Tyrannosaurid skeletal design first evolved at small body size. Science 326, 418–422. https://doi.org/10.1126/science.1177428 (2009).
Google Scholar
-
Currie, P. J. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontol. Pol. 48, 191–226. https://doi.org/10.7939/R3M90287B (2003).
Google Scholar
-
Voris, J. T. et al. Two exceptionally preserved juvenile specimens of Gorgosaurus libratus (Tyrannosauridae, Albertosaurinae) provide new insight into the timing of ontogenetic changes in tyrannosaurids. J. Vertebr. Paleontol. 41, e2041651. https://doi.org/10.1080/02724634.2021.2041651 (2022).
Google Scholar
-
Holtz, T., R., Jr. in Mesozoic Vertebrate life (eds K. Carpenter & D. Tanke) 64–83 (Indiana University Press, 2001).
-
Snively, E., Henderson, D. M. & Philips, D. S. Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics. Acta Palaeontol. Pol. 51, 435–454. https://doi.org/10.5281/zenodo.3739898 (2006).
Google Scholar
-
Tanke, D. H. & Currie, P. J. A history of Albertosaurus discoveries in Alberta, Canada. Can. J. Earth Sci. 47, 1197–1211. https://doi.org/10.1139/e10-057 (2010).
Google Scholar
-
Brusatte, S. L. The phylogeny of basal coelurosaurian theropods (Archosauria: Dinosauria) and patterns of morphological evolution during the dinosaur-bird transition Ph.D. thesis, Columbia University (2013).
-
Słowiak, J., Brusatte, S. L. & Szczygielski, T. Reassessment of the enigmatic Late Cretaceous theropod dinosaur, Bagaraatan ostromi. Zool. J. Linnean Soc. https://doi.org/10.1093/zoolinnean/zlad169 (2024).
Google Scholar
-
Brusatte, S. L., Benson, R. B. J. & Norell, M. A. The anatomy of Dryptosaurus aquilunguis (Dinosauria: Theropoda) and a review of its tyrannosauroid affinities. Am. Museum Novit. 8, 1–53. https://doi.org/10.1206/3717.2 (2011).
Google Scholar
-
Naish, D. & Cau, A. The osteology and affinities of Eotyrannus lengi, a tyrannosauroid theropod from the Wealden Supergroup of southern England. PeerJ 10, e12727. https://doi.org/10.7717/peerj.12727 (2022).
Google Scholar
-
Bakker, R. T., Williams, M. & Currie Philip, J. Nanotyrannus, a new genus of pygmy tyrannosaur, from the Latest Cretaceous of Montana. Hunteria 1, 1–30. https://doi.org/10.5281/zenodo.1037529 (1988).
Google Scholar
-
Fiorillo, A. R. & Tykoski, R. S. A diminutive new tyrannosaur from the top of the world. PLOS ONE 9, e91287. https://doi.org/10.1371/journal.pone.0091287 (2014).
Google Scholar
-
Carr, T. D. A high-resolution growth series of Tyrannosaurus rex obtained from multiple lines of evidence. PeerJ 8, e9192. https://doi.org/10.7717/peerj.9192 (2020).
Google Scholar
-
Druckenmiller, P. S., Erickson, G. M., Brinkman, D., Brown, C. M. & Eberle, J. J. Nesting at extreme polar latitudes by non-avian dinosaurs. Curr. Biol. 31, 3469–3478. https://doi.org/10.1016/j.cub.2021.05.041 (2021).
Google Scholar
-
Holtz, T. R. J. Theropod guild structure and the tyrannosaurid niche assimilation hypothesis: implications for predatory dinosaur macroecology and ontogeny in later Late Cretaceous Asiamerica1. Can. J. Earth Sci. 58, 778–795. https://doi.org/10.1139/cjes-2020-0174 (2021).
Google Scholar
-
Zhao, Q., Benton, M. J., Sullivan, C., Martin Sander, P. & Xu, X. Histology and postural change during the growth of the ceratopsian dinosaur Psittacosaurus lujiatunensis. Nat. Commun. https://doi.org/10.1038/ncomms3079 (2013).
Google Scholar
-
Carr, T. D., Varricchio, D. J., Sedlmayr, J. C., Roberts, E. M. & Moore, J. R. A new tyrannosaur with evidence for anagenesis and crocodile-like facial sensory system. Sci. Rep. 7, 44942. https://doi.org/10.1038/srep44942 (2017).
Google Scholar
-
Mesquite: a modular system for evolutionary analysis v. Version 3.81 (http://www.mesquiteproject.org, 2023).
-
Goloboff, P. A. & Morales, M. E. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39, 144–153. https://doi.org/10.1111/cla.12524 (2023).
Google Scholar
Acknowledgements
We thank Chaohe Yu (ZMNH) and Anhao Liu (ZMNH) for preparing the specimen, Shukang Zhang (IVPP) for helping to make thin sections, Qi Zhao (IVPP) and Te Guo (IVPP) for helping in the photograph of the thin section, Logan King (IVPP) for giving useful comments, which greatly improved the manuscript, and Ben Creisler for helping with etymological consultations. We thank Daniel Madzia and an anonymous reviewer for their constructive suggestions. Free online versions of TNT and Mesquite were provided by the Willi Hennig Society and the Free Software Foundation Inc.
Author information
Authors and Affiliations
Contributions
W.Z. performed the anatomical descriptive research, prepared the figures, analyzed the data, and wrote the paper. W.Z., X, J, J.X and T.D. reviewed, revised, and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Information.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Reprints and permissions
About this article
Cite this article
Zheng, W., Jin, X., Xie, J. et al. The first deep-snouted tyrannosaur from Upper Cretaceous Ganzhou City of southeastern China.
Sci Rep 14, 16276 (2024). https://doi.org/10.1038/s41598-024-66278-5
-
Received: 29 February 2024
-
Accepted: 01 July 2024
-
Published: 25 July 2024
-
DOI: https://doi.org/10.1038/s41598-024-66278-5
Keywords
Asiatyrannus xui
- Tyrannosauridae
- Nanxiong Formation
- Upper Cretaceous
- China
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.