Abstract
Before the rise of dinosaurs and pterosaurs, pseudosuchians—reptiles from the crocodilian lineage—dominated the Triassic land ecosystems. This lineage diversified into several less inclusive clades, resulting in a wide ecomorphological diversity during the Middle and Late Triassic. Some giant pseudosuchians occupied the top of the trophic webs, while others developed extensive bony armor as a defense mechanism, which later evolved as a convergence in the avemetatarsalian lineage. On the other hand, there were groups like the Gracilisuchidae, which was composed of carnivorous forms with lightweight build and less than 1 m in length. The fossil record of gracilisuchids is geographically restricted to China and Argentina, with one ambiguous record from Brazil. In the present study, the first unambiguous gracilisuchid from Brazil is described. Parvosuchus aurelioi gen. et sp. nov. comes from the Dinodontosaurus Assemblage Zone of the Santa Maria Formation, which is associated with the Ladinian-Carnian boundary. Composed of a complete cranium, vertebrae, pelvic girdle and hindlimbs, the new species nests with Gracilisuchus stipanicicorum and Maehary bonapartei in a phylogenetic analysis. Its discovery fills a taxonomic gap in Brazilian pseudosuchian fauna and reveals the smallest known member of this clade from the Dinodontosaurus Assemblage Zone, highlighting the diversity of pseudosuchians during the moment that preceded the dawn of dinosaurs.
Similar content being viewed by others

A new long-snouted marine reptile from the Middle Triassic of China illuminates pachypleurosauroid evolution

A new silesaurid from Carnian beds of Brazil fills a gap in the radiation of avian line archosaurs
A new desert-dwelling dinosaur (Theropoda, Noasaurinae) from the Cretaceous of south Brazil
Introduction
Shortly after the End-Permian extinction event1, the Triassic Period witnessed some of the most remarkable evolutionary episodes in the history of life, including the emergence of dinosaurs and pterosaurs2,3,4. However, reptiles from the crocodilian lineage—Pseudosuchia—ruled the land ecosystems before the dawn of the dinosaur age, achieving an impressive taxonomic and ecological diversity5. Some of the most impressive pseudosuchians from the Triassic Period were the giant quadrupedal apex predators, such as Prestosuchus chiniquensis6,7 and Luperosuchus fractus8. On the other hand, there were smaller faunivorous pseudosuchians that filled distinct niches, such as the lightly build gracilisuchids9. These small sized pseudosuchians were characterized by a relatively enlarged head with wide openings, carnivorous-like teeth, slender limbs, and a quadrupedal stance10,11,12,13,14. According to the fossil record, gracilisuchids lived from the Middle to the Late Triassic15,16. The oldest species is Turfanosuchus dabanensis15 and was recovered from the lower Kelamayi Formation of China, which is Anisian in age. A second species from China is Yonghesuchus sangbiensis12, which comes from the Upper part of Member II of the Tongchuan Formation, considered early Ladinian in age17. A putative coeval form comprises Gracilisuchus stipanicicorum10, from the Chañares Formation of Argentina18. Finally, a possible additional record from South America includes Maehary bonapartei16, which was excavated from the Caturrita Formation of Brazil (early Norian in age)19. This taxon was initially described as a pterosauromorph16. However, a recent phylogenetic study recovered it as a member of Gracilisuchidae2. If Maehary bonapartei is indeed a gracilisuchid, the fossil record of the group will extend to the early Norian. Despite this controversial record, no other specimen has been reported for Brazil20. This is particularly intriguing since the Triassic Brazilian beds are coeval with those in Argentina18, which have yielded records of gracilisuchids10. This gap is filled here with the description of the first unequivocal Brazilian member of this clade.
Material and methods
Institutional abbreviations
CAPPA/UFSM, Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia da Universidade Federal de Santa Maria, São João do Polêsine, Rio Grande do Sul, Brazil; MCP, Museu de Ciências e Tecnologia PUCRS, Porto Alegre, Rio Grande do Sul, Brazil.
Specimen
The specimen here described is housed at the Centro de Apoio à Pesquisa Paleontológica da Quarta Colônia/Universidade Federal de Santa Maria (CAPPA/UFSM), under the specimen number CAPPA/UFSM 0412.
Phylogenetic analysis
The phylogenetic affinities of the new pseudouchian archosaur were investigated employing the data matrix of Müller et al.2. The analysis was performed in the software TNT v. 1.521 following the parameters employed in the former study. Therefore, characters 9 and 119 were deactivated (these characters are non-independent from other characters added in previous iterations of this data matrix), all characters received the same weight, and characters 1, 2, 7, 10, 17, 19–21, 28, 29, 34, 36, 40, 42, 46, 50, 54, 66, 71, 74–76, 100, 122, 127, 146, 153, 156, 157, 171, 176, 177, 187, 202, 221, 227, 263, 266, 278, 279, 283, 324, 327, 331, 337, 342, 345, 351, 352, 354, 361, 365, 368, 370, 377, 379, 386, 387, 398, 410, 414, 424, 425, 430, 435, 446, 448, 454, 455, 458, 460, 463, 464, 470, 472, 478, 482, 483, 485, 489, 490, 502, 504, 510, 516, 520, 521, 529, 537, 546, 552, 556, 557, 567, 569, 571, 574, 581, 582, 588, 636, 648, 652, 662, 701, 731, 735, 737, 738, 743, 749, 766, 784, 803, 809, 810, 816, 850, 851, 872, 875, 885 and 888 were set as ordered. The final data matrix comprises 888 active characters and 215 active operational taxonomic units. Petrolacosaurus kansensis was used to root the most parsimonious trees (MPTs), which were recovered employing New Technology Search Algorithm, searching for a minimum length 100 times with the default Ratcheting, Drift, and Tree-fusing parameters. Consistency (CI) and retention (RI) indices were obtained employing the script by Spiekman et al.22 that does not take into account a priori deactivated terminals.
Nomenclatural acts
This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the International Code of Zoological Nomenclature. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix ‘http://zoobank.org/’. The LSID for this publication is: urn:lsid:zoobank.org:pub:80B862A6-E984-4AC1-801C-7100CC605BFF.
Results
Systematic paleontology
Archosauria Cope, 1869
Pseudosuchia Zittel, 1887–1890
Gracilisuchidae Butler et al., 2014
Parvosuchus aurelioi gen. et sp. nov.
[urn:lsid:zoobank.org:act:1CA45C46-8AD3-4515-9683-54A7423BE716 (genus)]
[urn:lsid:zoobank.org:act:0D35B9EA-FE83-4DBE-B1BD-AEF7E2CDC223 (species)]
Holotype
CAPPA/UFSM 0412, a partial skeleton, including a skull with lower jaws, 11 dorsal vertebrae, two sacral vertebrae, a complete pelvic girdle, both femora (lacking the distal portion), partial left tibia, partial left fibula, and left calcaneum.
Etymology
The genus name combines the Latin word “parvus” (= small) and the Greek word “suchus” (= crocodile). The specific epithet honors Pedro Lucas Porcela Aurélio for his passion for paleontology and prospecting, as well as for having discovered the fossil material described here.
Type locality, age, and horizon
Linha Várzea 2 (Becker) site (29°43′03″S, 53°09′07″W), municipalities of Paraíso do Sul, Rio Grande do Sul, Brazil (Fig. 1). This site belongs to the Pinheiros-Chiniquá Sequence23 of the Santa Maria Supersequence24, Paraná Basin. The fossiliferous content of this locality is associated with the Dinodontosaurus Assemblage Zone (AZ)25. Recent biostratigraphic studies suggest that this AZ has an age close to the Ladinian/Carnian boundary3,18,26.
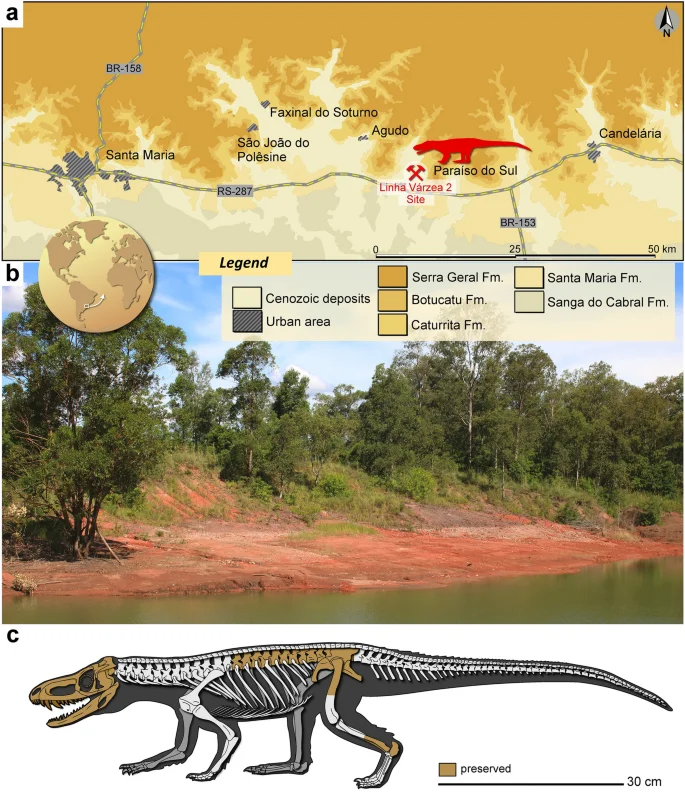
Provenance of Parvosuchus aurelioi gen. et sp. nov. (a) Location and geological context of the Linha Várzea 2 site, Paraíso do Sul, Rio Grande do Sul, Brazil. (b) General view of the Linha Várzea 2 site (taken in January 2023). (c) Hypothetical reconstruction of the skeleton of the Parvosuchus aurelioi gen. et sp. nov. depicting (in orange) the preserved portions of CAPPA/UFSM 0412. Unpreserved portions are based on the skeletal reconstruction of Gracilisuchus stipanicicorum by Jorge González14.
Diagnosis
Parvosuchus aurelioi differs from all other known gracilisuchids with comparable material in (*local autapomorphies): dorsal margin of the orbit markedly elevated above the skull table*; orbit dorsoventrally taller than long*; orbit shorter anteroposteriorly than the anteroposterior length of the antorbital fenestra; premaxilla lack a narial fossa; posterior portion of the parietal elevated above the skull roof*; parietal with a smooth anteromedial corner of the supratemporal fenestra; the ventral margin of the angular is directed upwards, forming an oblique angle with the ventral margin of the dentary*; craniomandibular joint above maxillary dental margin; and short pubis* with an anteroposterior distal expansion (see the “Discussion” for a differential diagnosis).
Description
Parvosuchus aurelioi was a relatively small pseudosuchian. Its skull (Fig. 2) is 144 mm in length (from the tip of the premaxilla to the occipital condyle). The premaxilla is anteroposteriorly short and bears an elongated posterolateral process that dorsally (Fig. 2a). The external naris is reduced and lack any associated fossa. A fossa occurs in the anteroventral corner of the external naris of Maehary bonapartei16. The number of tooth positions in the premaxilla is not clear. The alveolar margin of the premaxilla runs on the same plane that of the maxillary alveolar margin (Fig. 2b). The maxilla is elongated and gracile, with a posterior process that forms part of the antorbital fossa. Whereas there is a subtle dorsal expansion on the posterior portion of the posterior process, most of the posterior portion of this process presents a similar dorsoventral depth as the anterior portion ventral to the antorbital fenesta. A well-developed antorbital fossa excavated the ascending process of the maxilla. In lateral view, the ascending process present the width along all its length. Whereas the number of tooth positions is uncertain because of preservational biases, it is possible to note that the teeth reach the posterior portion of the bone. Some of the preserved teeth reveal a posteriorly recurved crown with a pointed apex. The poor preservation hampers the observation of possible serrations. The anterolateral process of the nasal is elongated and runs between the premaxilla and nasal. The bone forms a sharp lateral roof over the antorbital fenestra (Fig. 2c). The main body of the jugal is dorsoventrally tall and expanded transversely, forming a shelf at the ventral part of the orbital margin. There is a longitudinal ridge on the lateral surface of the bone (Fig. 2a,b), which is absent in Yonghesuchus sangbiensis12. The posterior process of the jugal is anteroposteriorly shorter than the main body, contrasting with Yonghesuchus sangbiensis12. The lacrimal forms the posterior portion of the dorsal margin of the antorbital fenestra. The orbit occupies approximately 17.3% of the total length of the skull, whereas the external antorbital fenestra comprises 33.7% of the length (Table 1). The orbit is taller than long, contrasting with the orbit of Gracilisuchus stipanicicorum and Turfanosuchus dabanensi anteroposteriorly, which is anteroposteriorly longer than tall9,10,11. The orbital margin of the frontal is dorsally projected, resulting in an orbital rim that strongly exceeds the dorsal margin of the skull roof. In Gracilisuchus stipanicicorum10 it is slightly elevated regarding the skull table, whereas in Turfanosuchus dabanensis11 it is not elevated. A small postorbital forms the posterodorsal corner of the orbital rim. The postorbital forms the main portion of the posterior border of the orbit. Distinct from Turfanosuchus dabanensis11, the ventral process if not distinctly anteriorly flexed. The squamosal folds over the quadrate head. The anterior portion of the ventral process contacts the posterior margin of the ventral process of the postorbital. As a result, the infratemporal fenestra is dorsoventrally short (Fig. 2a). This condition differs the specimen from the condition observed in Turfanosuchus dabanensis11, which bears an infratemporal fenestra that is divided in a ventral and dorsal fenestra by an anterior inflection of the squamosal. The supratemporal fenestra is transversely wider than it is anteroposteriorly long (Fig. 2d). The posterior portion of both parietals is elevated above the skull roof. There is a depression on the lateral surface of the main body of the parietal, which forms the medial margin of the supratemporal fossa. Although the bone surface is poorly preserved, it is possible to note that the dorsal surface of the skull is adorned with irregular sculptures on the nasal, frontal, and parietal bones. The holotype of Parvosuchus aurelioi preserves some ossicles within both orbital cavities, which are interpreted as parts of the slender sclerotic rings. The ring diameter is approximately 18 mm and the ossicles are approximately 2 mm in height.
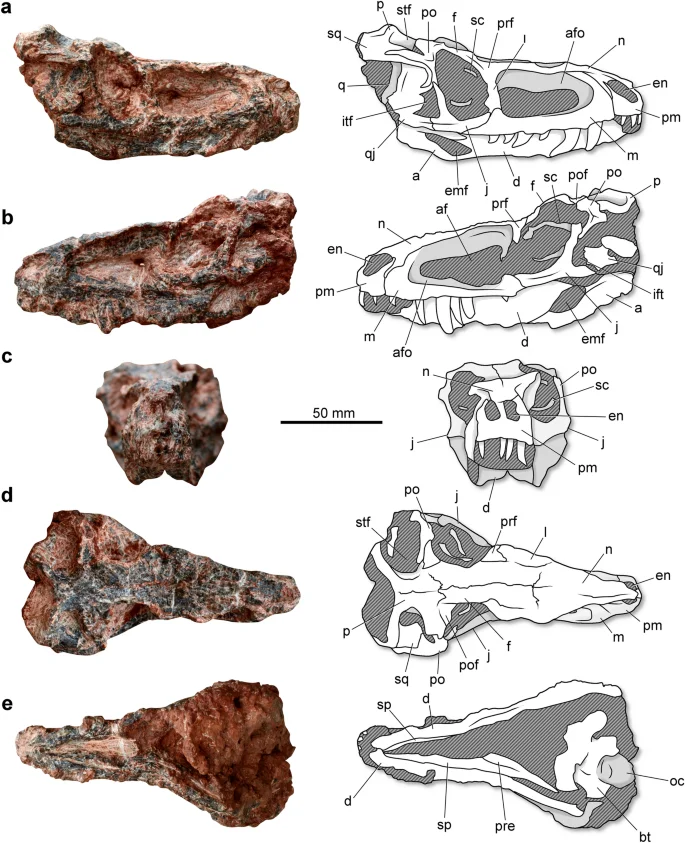
Skull and lower jaws of Parvosuchus aurelioi gen. et sp. nov. from the Pinheiros-Chiniquá Sequence (Ladinian-Carnian boundary) of the Santa Maria Supersequence, southern Brazil. Holotype (CAPPA/UFSM 0412) in right lateral (a), left lateral (b), anterior (c), dorsal (d), and ventral (e) views. a, angular; af, antorbital fenestra; afo, antorbital fossa; bt, basal tubera; d, dentary; emf, external mandibular fenestra; en, external naris; f, frontal; i, infratemporal fenestra; j, jugal; l, lacrimal; m, maxilla; n, nasal; o, occipital condyle; p, parietal; pm, premaxilla; po, postorbital; pof, postfrontal; pre, prearticular; prf, prefrontal; q, quadrato; qj, quadratojugal; sc, sclerotic ring; sp, splenial; sq, squamosal; stf, supratemporal fenestra.
The lower jaws are not entirely observable because the mandible is in occlusion. The mandibular symphysis is restricted to the anterior tip of the dentary, lacking any participation of the splenial (Fig. 2e). The slender dentary comprises the main portion of the lower jaw in lateral view. The bone reaches the level of the posterior half of the orbit and forms the anterior margin of the external mandibular fenestra (Fig. 2b). The posterior portion is composed of two processes. The ventral process is longer and contributes to the anterior half of the dorsal margin of the external mandibular fenestra. The presence of a surangular shelf is uncertain. In lateral view, the ventral margin of the angular is directed upwards (Fig. 2b), forming an oblique angle with the ventral margin of the dentary. In Gracilisuchus stipanicicorum10 and Yonghesuchus sangbiensis12 the ventral margin of the angular runs in the same longitudinal plane as the ventral margin of the dentary. The craniomandibular joint is just slightly above the maxillary dental margin.
The holotype of Parvosuchus aurelioi preserves a series of 13 articulated vertebrae, which includes 11 dorsal (6 to 16 dorsal elements) and the two sacral vertebrae (Fig. 3). The vertebral centra are spool-shaped and increase in length from the middle to the posterior elements (Table 2). There is shallow fossa that is not rimmed on the lateral surface of each centrum (Fig. 2a). The shape of the articular structures is poorly preserved. The anterior tip of the prezygapophysis barely extends beyond the anterior margin of their respective centrum. The parapophysis is situated on the neural arch in the observable elements. The transverse process is moderately expanded laterally. Neural spine of the posterior dorsal vertebrae is subrectangular and is slightly longer than tall. Furthermore, the spines lack a distal expansion. There are no neurocentral sutures. On the other hand, the bone surface of the specimen is poorly preserved. Therefore, this condition show be considered with carefully. The two sacral vertebrae are covered by a thick layer of concretion, hampering the observation of fine details. Nevertheless, it is safe to say that the sacrum is composed solely of two vertebrae (Fig. 4c). The neural spine of both vertebrae lacks a distal expansion. The sacral rib of the second vertebra expands anteroposteriorly at its distal end. The neural arch of the second sacral vertebra seems not fused to their respective centrum (Fig. 4d).

Axial skeleton of Parvosuchus aurelioi gen. et sp. nov. from the Pinheiros-Chiniquá Sequence (Ladinian-Carnian boundary) of the Santa Maria Supersequence, southern Brazil. Dorsal vertebrae of the holotype (CAPPA/UFSM 0412) in left lateral (a) and ventral (b) views. (c) Dorsal vertebrae 11 to 13 in left lateral view. ca, calcaneum; dp, diapophysis; dv, dorsal vertebra; fe, femur; fi, fibula; fo, fossa; il, ilium; is, ischium; ns, neural spine; poz, postzygapophysis; pp, parapophysis; prz, prezygapophysis; pu, pubis; sv, sacral vertebra.

Pelvic girdle and hindlimb of Parvosuchus aurelioi gen. et sp. nov. from the Pinheiros-Chiniquá Sequence (Ladinian-Carnian boundary) of the Santa Maria Supersequence, southern Brazil. Pelvic girdle of the holotype (CAPPA/UFSM 0412) in right lateral (a), left lateral (b), dorsal (c), and ventral (d) views. act, acetabulum; ca, calcaneum, cat, calcaneal tuber; dv, dorsal vertebra; fi, fibula; fe, femur; gt, greater trochanter of the femur; ib, iliac blade; is, ischium; isp ischiadic peduncle of the ilium; iss, ischial shaft; op, obturator plate of the ischium; poap, postacetabular process; prap, preacetabular process; pu, pubis; pup, pubic peduncle of the ilium; sac, supraacetabular crest; sv, sacral vertebra.
The ilium is short (Table 3), with approximately 53 mm in length. It comprises approximately 36.5% the total length of the skull. The preacetabular process is short and the anterior tip is rounded to sub-rectangular (Fig. 4b). There is no a vertical ridge posterior to the preacetabular process. The postacetabular process is elongated, with a posterior rounded to triangular margin (Fig. 4a). The main axis of the postacetabular process directs laterally along its posterior portion (Fig. 4c). The dorsal margin of the iliac blade is straight, such as in Turfanosuchus dabanensis12, whereas in Gracilisuchus stipanicicorum13 it is convex. The iliac blade is relatively low, as a result, the dorsal portion of the ilium is about the same height as the portion ventral to the supracetabular crest (i.e., acetabular region). The supracetabular crest rises from the lateral surface of the pubic peduncle and extends posteriorly, roofing the acetabulum. The crest smoothly merges with the main body of the bone dorsal the posterior half of the acetabulum. The ischiadic peduncle is short and lacks a posterior heel. In lateral view, the posterior transition between the ischiadic peduncle and the postacetabular process is concave. The medial wall of the iliac acetabulum is well-developed and convex ventrally.
The pubis is short (Table 3; approximately 43 mm in length) and anteroventrally oriented (Fig. 4b). There is a tuber on the anterolateral surface of the proximal portion. The iliac peduncle is anteroposteriorly elongated. The contribution of the pubis to the acetabulum is minimal do absent. The medial portion of the pubic apron is poorly preserved. There is a subtle anteroposterior expansion on the distal end of the pubis. The ischium is 41 mm in length (Table 3). The outline of the iliac peduncle is triangular and faces dorsally. An acetabular margin expands laterally (Fig. 4a). The contact with the pubis is not clear. The ischial shaft contacts its counterpart along its entire length. Conversely, there is no contact between the dorsal portion of the ischial shafts (Fig. 4d). As a result, there is a wide longitudinal groove between both ischia. The shaft is plate-like in cross-section. An obturator plate projects ventrally from the main body, extending along the shaft.
The total length of the hindlimb elements is uncertain because these bones are not entirely preserved. The proximal half of both femora is preserved. Conversely, there are no remains of the distal portion. The femoral head is slightly expanded in the anteroposterior axis (23 mm). It lacks any feature between the ventral transition from the head to the femoral shaft, resulting in a smooth transition (Fig. 4b). There is a straight and anteroposteriorly oriented groove on the proximal end, which is absent in Turfanosuchus dabanensis11. The anterolateral tuber is rounded and connects to a ridge that runs distally on the lateral surface of the femoral head. This longitudinal ridge bounds the anterior margin of a depression that extends along the posterior half of the lateral surface of the femoral head. The posteromedial tuber is well-developed. The proximal surface lacks any evidence of a trochanteric fossa. The greater trochanter is rounded. The proximal portion of the bone lacks an anterior (= lesser) trochanter and a trochanteric shelf. The mid-shaft is elliptical (Table 3), with the main axis anteroposteriorly oriented (13 mm axis anteroposterior and 9 mm axis lateromedial). The shape of the fourth trochanter is obscured by the matrix.
The specimen preserves the distal portions of the left tibia (Fig. 4a) and fibula (Fig. 4b), as well as the left calcaneum (Fig. 4b). The tibial shaft is stouter than that of the fibula. It is elliptical in cross-section. There is a longitudinal ridge on the anteromedial corner of the shaft. The tibia expands at its distal portion (Table 3), with the main axis of the expansion being anteroposteriorly oriented (14 mm). The anteromedial corner of the distal end of the tibia is rounded. In medial view, the anterior portion of the distal end is more distally projected than the posterior portion. Conversely, the posterior corner of the distal end expands posteriorly, resulting in a slightly concave posterior margin of the shaft. The fibular shaft is slender, expanding gradually towards the distal portion (Table 3). No ridges or tubers are present on the preserved portion of the shaft. The distal end of the fibula bends medially. The calcaneum is partially obscured by the fibula and ischia. The bone is anteroposteriorly elongated and bears a conspicuous calcaneal tuber. In lateral view, there is a marked notch between the calcaneal tuber and the calcaneal condyle. The latter forms the anterior portion of the bone. The posterior portion of the calcaneal tuber is upward directed. There is a fossa on the ventral/distal surface of the tuber.
Phylogenetic analysis
The analysis recovered 54,000 MPTs of 6773 steps each (CI = 0.17437; RI = 0.65586). In the strict consensus tree, Parvosuchus aurelioi is found as a member of Gracilisuchidae, within an unsolved node supporting Gracilisuchus stipanicicorum and Maehary bonapartei (Fig. 5a). This node is supported by: the ascending process of the maxilla with the same width along all its length in lateral view (ch. 60: 0 → 1); and posterior process of the maxilla with a similar dorsoventral depth as the anterior portion ventral to the antorbital fenestra (ch. 63: 0 → 1). Turfanosuchus dabanensis is the basalmost member of Gracilisuchidae. The clade is supported by 11 synapomorphies. Regarding the inner affinities of Gracilisuchidae, Yonghesuchus sangbiensis is the sister taxon of the node supporting Parvosuchus aurelioi, Gracilisuchus stipanicicorum and Maehary bonapartei. An interesting point comprises the position of Maehary bonapartei, which was not affected by the inclusion of Parvosuchus aurelioi. Therefore, the present analysis provides further support for including Maehary bonapartei in Gracilisuchidae2 rather than Pterosauromorpha16. Regarding the general topology recovered in the present analysis, the strict consensus tree maintains the same topology as that presented by Müller et al.2.

Results of the phylogenetic analysis and diversity of the Dinodontosaurus Assemblage Zone of Brazil. (a) Time-calibrated reduced strict consensus tree depicting the phylogenetic position of Parvosuchus aurelioi gen. et sp. nov. Number on nodes represent Bremer support values higher than 1. The temporal bars for each OTU represent the maximum and minimum ages of each geological unit. Divergence times set as approximately 1 million years. (b) Percentage of taxonomic groups recorded in the Dinodontosaurus Assemblage Zone according to the number of species. (c) Approximate body length of pseudosuchian species from the Dinodontosaurus ZA.
Discussion
Parvosuchus aurelioi is a lightly built pseudosuchian from the clade Gracilisuchidae. In addition to the phylogenetic analysis performed here, a series of typical traits of gracilisuchids are present in the holotype, reinforcing this assignation, such as: an enlarged antorbital fenestra; triangular expansion of the dorsal margin of the posterior process of the maxilla; expanded lateral margin of the nasal, forming a lateral roof; and size of the infratemporal fenestra is reduced due to the anterior expansion of the quadratojugal. Moreover, the new gracilisuchid differs from all the other members of the clade by a series of traits, as follows below.
Turfanosuchus dabanensis differs from Parvosuchus aurelioi in that the former has: dorsal margin of the orbit level with skull table or raised slightly; orbit dorsoventrally anteroposteriorly longer than tall; anterior portion of the nasal elevated above skull roof in lateral view; squamosal with an elongated ventral process; infratemporal fenestra divided into two fenestrae; posterior portion of the parietal not elevated above the skull roof; quadrate head partially exposed laterally; well-developed posteroventral process of the dentary; and longer pubis.
Gracilisuchus stipanicicorum differs from Parvosuchus aurelioi in that the former has: orbit anteroposteriorly longer than tall; orbit anteroposteriorly longer than the antorbital fenestra; slightly longer posterior process of the jugal; parietal extends over the interorbital region; parietal with a sinuous shelf on the anteromedial corner of the supratemporal fenestra; posterior portion of the parietal not elevated above the skull roof; ventral margin of the angular runs in the same longitudinal plane as the ventral margin of the dentary; craniomedial joint at the same level as the dental margin of the maxilla; convex dorsal margin of the iliac blade of the ilium; longer pubis; and pubis lack an anteroposterior distal expansion.
Yonghesuchus sangbiensis differs from Parvosuchus aurelioi in that the former has: elongated anterior portion of the maxilla; lack a ridge or bump on the lateral surface of the jugal; posterior process of the jugal is approximately half the total length of the bone (it is approximately 0.25 in Parvosuchus aurelioi); ventral margin of the angular runs in the same longitudinal plane as the ventral margin of the dentary; and craniomedial joint at the same level as the dental margin of the maxilla.
Maehary bonapartei differs from Parvosuchus aurelioi in that the former has: premaxilla with an expanded narial fossa in the anteroventral corner of the naris; lateral margin of the nasal is poorly developed, not forming a lateral shelf; and distal edge of the maxillary tooth crowns is straight or gently sigmoidin in labial or lingual views.
The inclusion of Parvosuchus aurelioi in the phylogenetic analysis did not affect the placement of Gracilisuchidae within the phylogenetic tree of Pseudosuchia, reinforcing the hypothesis that gracilisuchids are an early diverging clade of suchians. This same arrangement has been recovered in previous iterations of the present data matrix2,4,16,18,27, including through Bayesian phylogenetic analysis2,4. The present analysis also corroborates that gracilisuchids are more closely related to paracrocodylomorphs than to Ornithosuchidae, Erpetosuchidae, or Aetosauriforms.
Given that other gracilisuchids have not been identified within the Santa Maria Formation20, it is unlikely that Parvosuchus aurelioi represents a distinct ontogenetic stage of a known species. On the other hand, there is an enigmatic archosauromorph named Barberenasuchus brasiliensis28 from the same geological unit that yielded Parvosuchus aurelioi. Barberenasuchus brasiliensis is considered an indeterminate archosauriform with carnivorous feeding habits29. This archosauriform is particularly interesting because it is a small-sized animal (skull with less than 100 mm in length), representing one of the few records of a small reptile with predatory behavior from the Dinodontosaurus AZ. Conversely, the holotype (MCP-PV 220), the only known specimen, is poorly preserved, hampering properly systematic assignations29,30. Different interpretations have been proposed regarding the affinities of this taxon, such as a crocodylomorph31 or a dinosauromorph32. While it has not been classified as a gracilisuchid thus far, it is important to note that Barberenasuchus brasiliensis differs from Parvosuchus aurelioi in that the former has: dorsal margin of the orbit level with skull table; orbit anteroposteriorly longer than the antorbital fenestra; longer anterior process of the maxilla; dorsal end of the ascending process of the maxilla does not extend posteriorly to form the dorsal margin of the antorbital fossa; antorbital fossa has participation of the anterior process of the jugal; posterior margin of the dorsal portion of the parietal lies below the dorsal margin of the skull table; infratemporal is taller (taller than half of the orbital height); supratemporal fenestra anteroposteriorly longer than wide; lateral tip of the paraoccipital process exceed the lateral margin of the supratemporal fenestra in dorsal view; ventral margin of the angular runs in the same longitudinal plane as the ventral margin of the dentary; and craniomedial joint at the same level as the dental margin of the maxilla.
The Triassic ecosystems that preceded the radiation of dinosaurs were composed of a wide variety of reptiles from the crocodilian lineage18. This diversity is well documented in fossiliferous layers of the Chañares Formation in Argentina18 and Santa Maria Formation in Brazil20. New representatives from different groups of pseudosuchians have been reported in recent years from the Santa Maria Formation33,34. As a result, the diversity of pseudosuchian archosaurs has been increasing, but some groups that occur in the Chañares Formation had not yet been reported for Brazil. The discovery of Parvosuchus aurelioi fills the gap regarding the Gracilisuchidae. In addition to expanding the taxonomic diversity of pseudosuchians from the Dinodontosaurus AZ, the new species also provides important contributions to the understanding of the paleofaunal composition that existed in southern Brazil during the Ladinian to early Carnian. Represented by cynodonts35 and dicynodonts36, synapsids encompass the majority of the diversity within the Dinodontosaurus AZ20 (Fig. 5b). Whereas these dicynodonts are strictly large and herbivorous36, the record of cynodonts includes a certain ecomorphological diversity, encompassing carnivorous to omnivorous/herbivorous forms35. Far less abundant and diverse is the record of parareptiles, represented by the tiny Candelaria barbouri37,38, a owenettid procolophonoid. Archosauromorpha is relatively diverse, the clade is represented by herbivorous rhynchosaurus39, carnivorous proterochampsids40, and members of the clade Archosauria6,33,41,42. Actually, there is the problematic Barberenasuchus brasiliensis28,29,30, which was discussed above. Regarding the record of archosaurs, it is composed of both Pan-Aves and Pseudosuchia. Pan-Aves are represented by the aphanosaurian Spondylosoma absconditum43, the silesaur Gamatavus antiquus42, and an indeterminate dinosauromorph26. According to the holotype, Spondylosoma absconditum was carnivorous and reached approximately 1.75 m in length. Gamatavus antiquus was approximately 1.5 m in length and its feeding behaviors are uncertain. Whereas there are no teeth referred to this silesaur42, most members of this group are considered omnivorous or herbivorous44. Pseudosuchians are more diverse compared to Pan-Aves in the Dinodontosaurus AZ20. The clade is represented by Archeopelta arborensis45, Pagosvenator candelariensis41, Schultzsuchus loricarus34, Prestosuchus chiniquensis6,7, and Decuriasuchus quartacolonia33. Archeopelta arborensis and Pagosvenator candelariensis were carnivorous members of Erpetosuchidae18,27,41, both reached more than 2 m in length. Schultzsuchus loricarus was a large carnivorous from the clade Poposauroidea34. Finally, Prestosuchus chiniquensis and Decuriasuchus quartacolonia were carnivorous loricatans. Some recent histological analysis suggested that the specimens of Decuriasuchus quartacolonia were juvenile organisms46. Because these specimens are recorded from the same fossiliferous site that yielded specimens of Prestosuchus chiniquensis, it is plausible that they represent early ontogenetic stages of this large loricatan. Nevertheless, even the specimens of Decuriasuchus quartacolonia are not small animals, reaching approximately 2.5 m in length. The discovery of Parvosuchus aurelioi expands the diversity of pseudosuchians from the Dinodontosaurus AZ and represents the smallest pseudosuchian from this AZ (Fig. 5c). Bearing a skull with a length of 144 mm, Parvosuchus aurelioi is estimated to have reached approximately 1 m in body length based on comparisons with other related pseudosuchians. The inclusion of this new pseudosuchian in the paleofauna of the Dinodontosaurus AZ sheds light on the composition of terrestrial ecosystems that preceded the origin of dinosaurs in southern Brazil. The elongated and blade-like teeth of Parvosuchus aurelioi are typically associated with carnivorous feeding habits. Therefore, it represents the first small-sized predatory pseudosuchian from an environment dominated by huge archosaur predators (Fig. 6).

Artistic representation of a Middle-Late Triassic landscape of southern Brazil. (a) A large Prestosuchus chiniquensis feeds on the carcass of a dicynodont while individuals of Parvosuchus aurelioi gen. et sp. nov. compete for scraps. (b) and (c) depict details of individuals of Parvosuchus aurelioi gen. et sp. nov. Artwork by Matheus Fernandes.
The presence of a gracilisuchid in the Dinodontosaurus AZ reinforces the biostratigraphic similarity between this AZ and the Chañares Formation in Argentina. Nevertheless, the Chanãres Formation encompasses two distinct AZs, the Massetognathus-Chanaresuchus AZ and the Tarjadia AZ18. The Massetognathus-Chanaresuchus AZ overlaps the Tarjadia AZ and is early Carnian in age according to radioisotopic datings47, whereas the Tarjadia AZ is assigned to the Ladinian-Carnian boundary18. The shared presence of phylogenetic related erpetosuchids, rhynchosaurs, suchians, and cynodonts between the Massetognathus-Chanaresuchus AZ and the Dinodontosaurus AZ provided support for biostratigraphic proposals correlating both AZs3,18,20,26. On the other hand, the record of some groups of cynodonts35, dinosauromorphs42, and proterochampsids40 in the Dinodontosaurus AZ resembles more the fossil content of the Massetognathus-Chanaresuchus AZ. Similarly, whereas gracilisuchids are absent in the Tarjadia AZ, the clade occurs in the Massetognathus-Chanaresuchus AZ18. Moreover, radioisotopic datings suggested a maximum depositional age of 237 Ma (early Carnian) to the Brazilian Santacruzodon AZ48. This AZ overlaps the Dinodontosaurus AZ20,23, suggesting that it is more likely associated with the Ladinian-Carnian boundary or even older. Therefore, the occurrence of the gracilisuchid Parvosuchus aurelioi in the Dinodontosaurus AZ emphasizes the need for refinement in recognizing the different paleofaunas commonly assigned to the Dinodontosaurus AZ.
Conclusions
The new specimen described here bears a unique set of traits that led to the naming of a new pesudosuchian taxon. Parvosuchus aurelioi is the first Brazilian gracilisuchid found in layers attributed to the Ladinian-Carnian boundary. The inclusion of a small carnivorous pseudosuchian in the paleofauna of the Dinodontosaurus AZ expands our knowledge regarding the faunal composition of this biostratigraphic unit. Lastly, the occurrence of gracilisuchids in the same AZ that contains large paracrocodylomorphs, erpetosuchids, and rhynchosaurs demonstrates a complex biostratigraphic context, suggesting that the Dinodontosaurus AZ may contain subdivisions that have not yet been recognized.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
-
Benton, M. J. & Twitchett, R. J. How to kill (almost) all life: The end-Permian extinction event. Trends Ecol. Evol. 18, 358–365 (2003).
Google Scholar
-
Müller, R. T. et al. New reptile shows dinosaurs and pterosaurs evolved among diverse precursors. Nature 620, 589–594 (2023).
Google Scholar
-
Novas, F. E. et al. Review of the fossil record of early dinosaurs from South America, and its phylogenetic implications. J. S. Am. Earth Sci. 110, 103341 (2021).
Google Scholar
-
Ezcurra, M. D. et al. Enigmatic dinosaur precursors bridge the gap to the origin of Pterosauria. Nature 588, 445–449 (2020).
Google Scholar
-
Foth, C., Ezcurra, M. D., Sookias, R. B., Brusatte, S. L. & Butler, R. J. Unappreciated diversification of stem archosaurs during the Middle Triassic predated the dominance of dinosaurs. BMC Evol. Biol. 16, 1–10 (2016).
Google Scholar
-
Roberto-Da-Silva, L., Müller, R. T., de França, M. A. G., Cabreira, S. F. & Dias-Da-Silva, S. An impressive skeleton of the giant top predator Prestosuchus chiniquensis (Pseudosuchia: Loricata) from the Triassic of Southern Brazil, with phylogenetic remarks. Hist. Biol. 32, 976–995 (2018).
Google Scholar
-
Desojo, J. B., Von Baczko, B. & Rauhut, O. W. M. Anatomy, taxonomy and phylogenetic relationships of Prestosuchus chiniquensis (Archosauria: Pseudosuchia) from the original collection of von Huene, Middle-Late Triassic of southern Brazil. Palaeontol. Electron. 23, a04 (2020).
-
Desojo, J. B. & Arcucci, A. B. New material of Luperosuchus fractus (Archosauria: Crurotarsi) from the Middle Triassic of Argentina: The earliest known South American ‘rauisuchian’. J. Vertebr. Paleontol. 29, 1311–1315 (2009).
Google Scholar
-
Butler, R. J. et al. New clade of enigmatic early archosaurs yields insights into early pseudosuchian phylogeny and the biogeography of the archosaur radiation. BMC Evol. Biol. 14, 1–16 (2014).
Google Scholar
-
Romer, A. S. The Chañares (Argentina) Triassic reptile fauna. An early ornithosuchid pseudosuchian, Gracilisuchus stipanicicorum, gen. et sp. nov. Breviora 389, 1–24 (1972).
-
Wu, X. C. & Russell, A. P. Redescription of Turfanosuchus dabanensis (Archosauriformes) and new information on its phylogenetic relationships. J. Vertebr. Paleontol. 21, 40–50 (2001).
Google Scholar
-
Wu, X. C., Liu, J. & Li, J.-L. The anatomy of the first archosauriform (Diapsida) from the terrestrial Upper Triassic of China. Vertebrata PalAsiatica 39, 251–265 (2001).
-
Lecuona, A. & Desojo, J. B. Hind limb osteology of Gracilisuchus stipanicicorum (Archosauria: Pseudosuchia). Earth Environ. Sci. Trans. R. Soc. Edinb. 102, 105–128 (2012).
-
Lecuona, A., Desojo, J. B. & Pol, D. New information on the postcranial skeleton of Gracilisuchus stipanicicorum (Archosauria: Suchia) and reappraisal of its phylogenetic position. Zool. J. Linn. Soc. 181, 638–677 (2017).
Google Scholar
-
Young, C. C. On a new pseudosuchian from Turfan, Sinking (Xinjiang). Mem. Inst. Vertebr. Paleontol. Paleoanthropol. Acad. Sin. Ser. B 10, 15–37 (1973).
-
Kellner, A. W. et al. Reassessment of Faxinalipterus minimus, a purported Triassic pterosaur from southern Brazil with the description of a new taxon. PeerJ 10, e13276 (2022).
Google Scholar
-
Liu, J. et al. High-precision temporal calibration of Middle Triassic vertebrate biostratigraphy: U-Pb zircon constraints for the Sinokannemeyeria Fauna and Yonghesuchus. Vertebr. PalAsiatica 56, 16–24 (2018).
-
Ezcurra, M. D. et al. Deep faunistic turnovers preceded the rise of dinosaurs in southwestern Pangaea. Nat. Ecol. Evol. 1(1477–1483), 8 (2017).
-
Langer, M. C., Ramezani, J. & Da-Rosa, Á. A. S. Upb age constraints on dinosaur rise from south Brazil. Gondwana Res. 57, 133–140 (2018).
Google Scholar
-
Schultz, C. L. et al. Triassic faunal successions of the Paraná Basin, southern Brazil. J. S. Am. Earth Sci. 104, 102846 (2020).
Google Scholar
-
Goloboff, P. A. & Catalano, S. A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32, 221–238 (2016).
Google Scholar
-
Spiekman, S. N., Ezcurra, M. D., Butler, R. J., Fraser, N. C. & Maidment, S. C. Pendraig milnerae, a new small-sized coelophysoid theropod from the Late Triassic of Wales. R. Soc. Open Sci. 8, 210915 (2021).
Google Scholar
-
Horn, B. L. D. et al. A new third-order sequence stratigraphic framework applied to the Triassic of the Paraná Basin, Rio Grande do Sul, Brazil, based on structural, stratigraphic and paleontological data. J. S. Am. Earth Sci. 55, 123–132 (2014).
Google Scholar
-
Zerfass, H. et al. Sequence stratigraphy of continental Triassic strata of Southernmost Brazil: A contribution to southwestern Gondwana palaeogeography and palaeoclimate. Sediment. Geol. 161, 85–105 (2003).
Google Scholar
-
Da-Rosa, Á. A. S., Schwanke, C., Aurélio, P. L. P., Poitevin, M. & Neto, L. W. Sítio Linha Várzea-Uma nova assembléia fossilífera do Triássico Médio do Sul do Brasil. Geociências 2, 115–129 (2005).
-
Müller, R. T. & Garcia, M. S. Oldest dinosauromorph from South America and the early radiation of dinosaur precursors in Gondwana. Gondwana Res. 107, 42–48 (2022).
Google Scholar
-
Müller, R. T., Belen von Baczko, M., Desojo, J. B. & Nesbitt, S. J. The first ornithosuchid from Brazil and its macroevolutionary and phylogenetic implications for Late Triassic faunas in Gondwana. Acta Palaeontol. Polonica 65, 1–10 (2020).
Google Scholar
-
Mattar, L. C. B. Sobre Barberenasuchus brasiliensis gen. et sp. nov., do Mesotriássico do Rio Grande do Sul. I. Diagnoses genérica e específica. Anais do Congresso Brasileiro de Paleontologia 10, 77–83 (1987).
-
Irmis, R. B., Nesbitt, S. J. & Sues, H.-D. Early Crocodylomorpha. Geol. Soc. Lond. Special Publ. 379, 275–302 (2013).
Google Scholar
-
Leardi, J. M., Yáñez, I. & Pol, D. South American Crocodylomorphs (Archosauria; Crocodylomorpha): A review of the early fossil record in the continent and its relevance on understanding the origins of the clade. J. S. Am. Earth Sci. 104, 102780 (2020).
Google Scholar
-
Walker, A. D. A revision of the Jurassic reptile Hallopus victor (Marsh) with remarks on the classification of the crocodiles. Philos. Trans. R. Soc. Lond. B 257, 323–372 (1970).
Google Scholar
-
França, M. A., Bittencourt, J. & Langer, M. C. Reavaliação taxonômica de Barberenasuchus brasiliensis (Archosauriformes), Ladiniano do Rio Grande do Sul (Zona-Assembléia de Dinodontosaurus). Paleontologia em Destaque 28, 230 (2013).
-
França, M. A. G., Ferigolo, J. & Langer, M. C. Associated skeletons of a new middle Triassic “Rauisuchia” from Brazil. Naturwissenschaften 98, 389–395 (2011).
Google Scholar
-
Desojo, J. B. & Rauhut, O. W. Reassessment of the enigmatic “Prestosuchus” loricatus (Archosauria: Pseudosuchia) from the Middle-Late Triassic of southern Brazil. The Anatomical Record, Online First (2024).
-
Martinelli, A. G. et al. The African cynodont Aleodon (Cynodontia, Probainognathia) in the Triassic of southern Brazil and its biostratigraphic significance. PLoS ONE 12, e0177948 (2017).
Google Scholar
-
Kammerer, C. F. & de los Angeles Ordonez, M. Dicynodonts (Therapsida: Anomodontia) of South America. J. S. Am. Earth Sci. 108, 103171 (2021).
Google Scholar
-
Price, L. I. Um procolofonídeo do Triássico do Rio Grande do Sul. Boletim da Divisao de Geologia e Mineralogia 122, 7–27 (1947).
-
Cisneros, J. C. et al. A procolophonoid reptile with temporal fenestration from the Middle Triassic of Brazil. Proc. R. Soc. Lond. Ser. B 271, 1541–1546 (2004).
Google Scholar
-
Schultz, C. L., Langer, M. C. & Montefeltro, F. C. A new rhynchosaur from south Brazil (Santa Maria Formation) and rhynchosaur diversity patterns across the Middle-Late Triassic boundary. PalZ 90, 593–609 (2016).
Google Scholar
-
Paes-Neto, V. D. et al. New rhadinosuchine proterochampsids from the late Middle‐early Late Triassic of southern Brazil enhance the diversity of archosauriforms. The Anatomical Record, Online First (2023).
-
Lacerda, M. B., De Franca, M. A. & Schultz, C. L. A new erpetosuchid (Pseudosuchia, Archosauria) from the middle–late Triassic of southern Brazil. Zool. J. Linn. Soc. 184, 804–824 (2018).
Google Scholar
-
Pretto, F. A. et al. The oldest South American silesaurid: New remains from the Middle Triassic (Pinheiros-Chiniquá Sequence, Dinodontosaurus Assemblage Zone) increase the time range of silesaurid fossil record in southern Brazil. J. S. Am. Earth Sci. 120, 104039 (2022).
Google Scholar
-
Nesbitt, S. J. et al. The earliest bird-line archosaurs and the assembly of the dinosaur body plan. Nature 544, 484–487 (2017).
Google Scholar
-
Müller, R. T. & Garcia, M. S. A paraphyletic ‘Silesauridae’ as an alternative hypothesis for the initial radiation of ornithischian dinosaurs. Biol. Lett. 16, 20200417 (2020).
Google Scholar
-
Desojo, J. B., Ezcurra, M. D. & Schultz, C. L. An unusual new archosauriform from the Middle-Late Triassic of southern Brazil and the monophyly of Doswelliidae. Zool. J. Linn. Soc. 161, 839–871 (2011).
Google Scholar
-
Farias, B. D. M. D. et al. Bone histology supports gregarious behavior and an early ontogenetic stage to Decuriasuchus quartacolonia (Pseudosuchia: Loricata) from the Middle‐Late Triassic of Brazil. The Anatomical Record. Online First (2023).
-
Marsicano, C. A., Irmis, R. B., Mancuso, A. C., Mundil, R. & Chemale, F. The precise temporal calibration of dinosaur origins. Proc. Natl. Acad. Sci. 113, 509–513 (2016).
Google Scholar
-
Philipp, R. P. et al. Middle Triassic SW Gondwana paleogeography and sedimentary dispersal revealed by integration of stratigraphy and U-Pb zircon analysis: the Santa Cruz Sequence, Paraná Basin, Brazil. J. S. Am. Earth Sci. 88, 216–237 (2018).
Google Scholar
Acknowledgements
I am deeply grateful to Pedro Lucas Porcela Aurélio for the discovery of the holotype described here. I extend my gratitude to the two anonymous reviewers for their valuable comments and suggestions, which greatly improved the manuscript. Access to the free version of TNT 1.5 was possible owing to the Willi Hennig Society. This work was carried out with aid of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 404095/2021-6; 303034/2022-0; and 406902/2022-4).
Author information
Authors and Affiliations
Contributions
R.T.M. data curation, funding acquisition, formal analysis, investigation, writing.
Corresponding author
Ethics declarations
Competing interests
The author declares no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Supplementary Information.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
Reprints and permissions
About this article
Cite this article
Müller, R.T. A new small-sized predatory pseudosuchian archosaur from the Middle-Late Triassic of Southern Brazil.
Sci Rep 14, 12706 (2024). https://doi.org/10.1038/s41598-024-63313-3
-
Received: 26 February 2024
-
Accepted: 27 May 2024
-
Published: 20 June 2024
-
DOI: https://doi.org/10.1038/s41598-024-63313-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.