Abstract
We compared the protective effects of inactivated SARS-CoV-2 vaccines derived from the ancestral and the currently circulating BA.5.2 strains against infection with multiple variants in Syrian golden hamsters. Vaccination with BA.5.2 effectively protected against infection with the Omicron subvariants including XBB.1, but not the Alpha or Delta variant. In contrast, hamsters vaccinated with the ancestral strain demonstrated decent neutralization activity against both the Omicron and non-Omicron variants. Our findings might instruct future design and formulation of SARS-CoV-2 vaccines.
Vaccination against SARS-CoV-2 has been very effective in the protection against severe disease and death, but it was less successful in the prevention of viral transmission1,2. To deal with the emergence of SARS-CoV-2 variants of concern (VOCs) that exhibit increased immune evasiveness, viral strains used for vaccine production need to be updated periodically3. Whereas BA.2 and BA.5 subvariants of SARS-CoV-2 Omicron were consecutively predominant worldwide in early and mid-2022, they gave way to other subvariants such as BQ.1, BQ.1.1, XBB.1, XBB.1.5 and BF.7 in some parts of the world, but BA.5 and BA.2 continue to dominate in other areas2,4. One future direction is to formulate one single type of vaccine for annual booster immunization against the circulating strains, coupled to influenza vaccination3. However, questions remain unanswered as to whether a monovalent or bivalent SARS-CoV-2 vaccine should be used and whether a monovalent vaccine against the predominant strain could have effective protection against sufficiently broad range of VOCs. To shed light on these questions, in this study we asked whether vaccination of Syrian golden hamsters with inactivated SARS-CoV-2 Omicron BA.5.2 subvariant might provide good protection against emerging subvariants, such as XBB.1, and VOCs that circulated before the arrival of the Omicron variant. Side-by-side comparison with the inactivated vaccine derived from the ancestral strain was made.
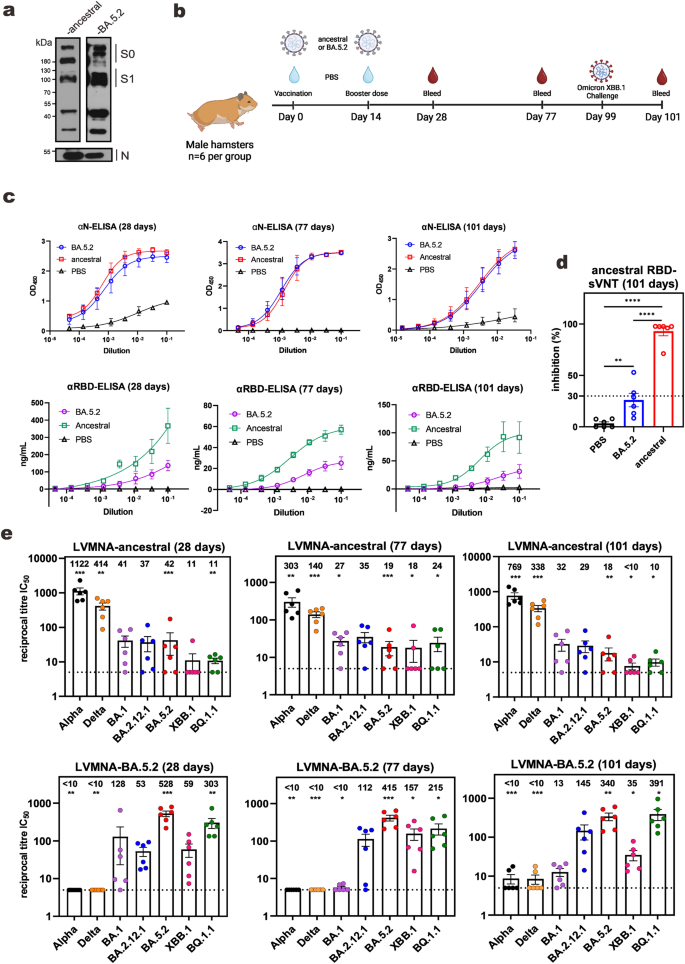
SARS-CoV-2 variants propagated in VeroE6-TMPRSS2 cells for 5 days were inactivated. As verified by immunoblotting, the purified virions of the ancestral (Wuhan-1) and the Omicron BA.5.2 strains of SARS-CoV-2 contained abundant viral spike (S) and N proteins (Fig. 1a, Supplementary Fig. 1). Syrian golden hamsters (6 per group) were vaccinated twice with 3 μg each of inactivated virions of BA.5.2 or ancestral strains in a prime-boost scheme with an interval of 14 days (Fig. 1b). Anti-N antibodies were equally robust in animals vaccinated with BA.5.2 and ancestral strains after 28, 77, and 101 days (Fig. 1c). In stark contrast, the levels of anti-RBD of the ancestral strain were considerably lower in BA.5.2-vaccinated hamsters than in animals vaccinated with the ancestral strain at all three time points. Consistently, sera from BA.5.2-vaccinated hamsters displayed much less pronounced inhibitory activity on the binding of ACE2 to the RBD of the ancestral strain in the surrogate virus neutralization test (sVNT) (Fig. 1d). Furthermore, when live virus microneutralization assay (LVMNA) was carried out with multiple SARS-CoV-2 VOCs, sera of BA.5.2-vaccinated hamsters exhibited strong neutralizing activity against BA.5.2 as well as other Omicron subvariants BA.1, BA.2.12.1, BQ.1.1 and XBB.1 at 28 days post-vaccination (Fig. 1e, Supplementary Fig. 2). The neutralizing activity against BA.1 decreased to right above basal after 77 days, when the neutralization of BA.5.2, BA.2.12.1, BQ.1.1 and XBB.1 remained robust. However, these sera had almost lost the ability to neutralize Alpha or Beta strain at the same time point, with only 2 out of 6 sera capable of neutralizing. Opposite to this pattern, sera from animals vaccinated with the ancestral strain effectively neutralized the Alpha and Delta variants at both early and late time points. Moreover, their neutralizing activity against BA.1, BA2.12.1 and BA.5 was well above the basal level. However, the sera could marginally neutralize XBB.1 (Fig. 1e). Interestingly, the protective efficacy differences between ancestral- and BA.5.2-vaccinated groups become increasingly prominent as time progresses.

a Immunoblot for proteins in SARS-CoV-2 virions purified by sucrose-gradient centrifugation for vaccine preparation. The uncleaved S0 and the cleaved S1 are indicated. All virus stocks have been verified by Nanopore sequencing after passaging in VeroE6-TMPRSS2 cells. Rabbit monoclonal anti-RBD is from ThermoFisher (HL257). Mouse monoclonal anti-N was home-made. b Vaccination scheme of hamsters. Schematic diagram was created using BioRender.com. c ELISA assays. Hamster sera were collected at 28, 77, and 101 days post-vaccination for analysis of anti-RBD (αRBD) or anti-N (αN). For anti-RBD, a standard curve plotted from known concentrations of an anti-SARS-CoV-2 RBD monoclonal antibody (HL257, MA5-26353, ThermoFisher Scientific) was used to determine antibody concentration. Results are from three independent experiments. Error bars represent mean ± standard deviation (SD). d sVNT assays. Error bars represent mean ± standard error mean (SEM) (n = 6). e LVMNA assays. Limit of detection (LOD) is 5 (dotted line). Error bars represent mean ± SEM (n = 6). *: P < 0.05. **: P < 0.01. ***: P < 0.001. ****: P < 0.0001. IC50: 50% inhibitory concentration.
When challenged with XBB.1, significantly lower numbers of SARS-CoV-2 sub-genomic envelop (E) gene copies were found in both lung and tracheal tissues of vaccinated hamsters (Fig. 2a). However, there was no significant inter-group difference in nasal wash samples. It remained to be clarified whether residual and dead RNA might persist in the nasal cavity. Thus, we went on to perform plaque assays in parellel to quantify infectious virions in the samples. As expected, amounts of infectious virions in lung, tracheal and nasal wash samples of the BA.5.2-vaccinated animals, as measured by plaque assays, were remarkably lower (Fig. 2b). This reduction was less pronounced in lung and trachea of animals vaccinated with the ancestral strain. In addition, mRNA expression levels of proinflammatory cytokines interleukin-4 (IL-4) and IL-6 as well as chemokine CCL-17 increased in the lung of BA.5.2-vaccinated hamsters (Fig. 2c). Histopathological analysis revealed minimal immune cell infiltration and alveolar wall edema in the lung of these animals. Histological scoring of the lung sections reveals significant severity in the PBS-treated hamsters, but not vaccinated hamsters (Fig. 2d).

a Subgenomic E gene copies in respiratory samples. Hamsters were vaccinated and challenged with XBB.1 at 99 days post-vaccination. Samples were collected for RT-qPCR analysis at 2 dpi. One-way ANOVA was performed to determine significant differences between groups. Error bars represent mean ± SEM (n = 6). b Viral titers in respiratory samples at 2 dpi by plaque assays. The limit of detection (LOD) is 100. One-way ANOVA was performed to determine significant difference between groups. Error bars represent mean ± SEM. c Cytokine and chemokine profiling of the lung at 2 dpi by RT-qPCR. One-way ANOVA was performed to determine significant difference between groups. Error bars represent mean ± SEM (n = 6). d Hematoxylin and eosin staining of the lung at 2 dpi. Histological scores were derived as described15. Scale bar: 100 µm. One-way ANOVA was performed to determine significant difference between groups. Error bars represent mean ± SEM (n = 8). *: P < 0.05. **: P < 0.01. ***: P < 0.001. ns not significant (P > 0.05). pfu plaque forming unit. au arbitrary unit. HKG house-keeping gene (β-actin).
Whether a monovalent or bivalent vaccine should be used for post-pandemic vaccination against SARS-CoV-2 remains to be understood5. Bivalent mRNA vaccines expressing S of both ancestral and BA.5 strains induce broadly neutralizing antibodies against existing VOCs in animals and humans6,7, providing good protection against symptomatic infection with BA.5 and XBB.1 subvariants. Our demonstration of the cross-protection of Omicron subvariants by vaccination with inactivated BA.5 is generally consistent with this notion. Plausibly, vaccination with BA.5 should provide long-lasting protection against other Omicron subvariants, including emerging subvariants and subvariants to be emerged. It will be of interest to see whether the bivalent vaccine could be replaced by one single monovalent vaccine directed against the predominant strain currently circulating in the world.
Our finding that vaccination with inactivated BA.5 was less effective in protecting against non-Omicron variants suggests that inclusion of a non-Omicron strain in the SARS-CoV-2 vaccine might be desirable5,8. Most people in the general population were infected by the Omicron variant in 2022, but much less might have been exposed to the non-Omicron variants9. Particularly, some children and a small subset of adults might still be both unvaccinated and uninfected. Whereas the Delta variant has been found to be highly fit in immunologically naïve population10, the Omicron variant might have outcompeted the Delta variant only in vaccinated and/or infected people11. In other words, pre-existing immunity against non-Omicron variants in humans could have driven the evolution of the Omicron, which is not only most diverged from the non-Omicron variants but also most immune evasive1,2. It will be intriguing to investigate whether the absence of a non-Omicron variant in the SARS-CoV-2 vaccine could facilitate the re-selection and re-emergence of the Delta variant, which is more pathogenic than Omicron.
Exclusion from the composition of the updated SARS-CoV-2 vaccine of the ancestral strain that is no longer circulating has been proposed3. Before we can reach a conclusion on this, several important questions must be answered. First, whether previously circulated strains of SARS-CoV-2 could re-emerge and re-circulate as those of influenza viruses remains to be seen. Second, whether strong pre-existing immunity against non-Omicron variants acquired through vaccination and/or natural infection might guard against the re-emergence of the non-Omicron variants as suggested above or emergence of another Delta-like variant should be clarified. Finally, whether the pre-existing immunity against non-Omicron variants would be long-lasting and even enhanced via booster vaccination and/or natural infection with an Omicron subvariant merits further investigations12. Nevertheless, inclusion of a non-Omicron strain in the SARS-CoV-2 vaccine might not be unwise.
Methods
Viruses
SARS-CoV-2 Omicron XBB.1 variant was isolated from respiratory tract specimens of laboratory-confirmed COVID-19 patient in Hong Kong. Other clinical isolates of SARS-CoV-2 have been described previously13,14. VeroE6-TMPRSS2 cells were cultured until confluency. Viruses were inoculated the next day and incubated at 37 °C for 5 days. Viral supernatant was harvested and inactivated with paraformaldehyde to a final concentration of 0.2% at 4 °C for 5 more days. Virions were concentrated and purified as described5. Purified inactivated SARS-CoV-2 containing 3 μg of protein was mixed at 1:1 ratio with ImjectTM Alum Adjuvant (Thermo Scientific, 77161) before injected intramuscularly into Syrian golden hamsters. A booster dose was given after 14 days.
ELISA assays for SARS-CoV-2 spike receptor binding domain (RBD) or nucleocapsid, surrogate virus neutralization test (sVNT) using cPassTM SARS-CoV-2 Neutralization Antibody Detection Kit, and live virus microneutralization assay (LVMNA) were carried out as previously described15.
Infection of Syrian golden hamsters
All protocols for animal experiments have been approved by the Committee on the Use of Live Animals in Teaching and Research (CULATR) of the University of Hong Kong (Reference code: CULATR 5754-21). They were also performed in accordance with the standard operating procedures of Biosafety Level 3 animal facilities. Male Syrian golden hamsters aged 4-6 weeks old were used. At day 99 post-vaccination, hamsters anesthetized with ketamine (200 mg/kg) and xylazine (10 mg/kg) intraperitoneally were intranasally challenged with 105 PFU (in 50 μL) of SARS-CoV-2 Omicron XBB.1 subvariant. At 2 days post-infection (dpi), hamsters were sacrificed via intraperitoneal injection of dorminal (200 μL) for virological and histopathological evaluation. RT-qPCR and Western blotting were performed as described5,15. Primers used are listed in Supplementary Table 1.
Statistical analysis
Statistical significance was assessed by unpaired two-tailed Student’s t test or one-way analysis of variance (ANOVA) with Welch’s correction and Dunnett’s post-hoc test using Prism version 8.4.3.
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Data supporting the findings of this study are available in this paper, in the Supplementary Information, or from the corresponding author upon reasonable request.
References
-
Barouch, D. H. COVID-19 vaccines – Immunity, variants, boosters. N. Engl. J. Med. 387, 1011–1020 (2022).
Google Scholar
-
Carabelli, A. M. et al. SARS-CoV-2 variant biology: immune escape, transmission and fitness. Nat. Rev. Microbiol. 21, 162–177 (2023).
Google Scholar
-
Collier, A. Y. et al. Immunogenicity of BA.5 bivalent mRNA vaccine boosters. N. Engl. J. Med. 388, 565–567 (2023).
Google Scholar
-
Velavan, T. P., Ntoumi, F., Kremsner, P. G., Lee, S. S. & Meyer, C. G. Emergence and geographic dominance of Omicron subvariants XBB/XBB.1.5 and BF.7 – the public health challenges. Int. J. Infect. Dis. 128, 307–309 (2023).
Google Scholar
-
Ye, Z. W. et al. Cross-variant protection against SARS-CoV-2 infection in hamsters immunized with monovalent and bivalent inactivated vaccines. Int. J. Biol. Sci. 18, 4781–4791 (2022).
Google Scholar
-
Link-Gelles, R. et al. Early estimates of bivalent mRNA booster dose vaccine effectiveness in preventing symptomatic SARS-CoV-2 infection attributable to Omicron BA.5- and XBB/XBB.1.5-related sublineages among immunocompetent adults—increasing community access to testing program, United States, December 2022-January 2023. Morb. Mortal. Wkly Rep. 72, 119–124 (2023).
Google Scholar
-
Muik, A. et al. Exposure to BA.4/5 S protein drives neutralization of Omicron BA.1, BA.2, BA.2.12.1, and BA.4/5 in vaccine-experienced humans and mice. Sci. Immunol. 7, eade9888 (2022).
Google Scholar
-
Tseng, H. F. et al. Effectiveness of mRNA-1273 vaccination against SARS-CoV-2 omicron subvariants BA.1, BA.2, BA.2.12.1, BA.4, and BA.5. Nat. Commun. 14, 189 (2023).
Google Scholar
-
Erikstrup, C. et al. Seroprevalence and infection fatality rate of the SARS-CoV-2 Omicron variant in Denmark: A nationwide serosurveillance study. Lancet Reg. Health Eur. 21, 100479 (2022).
Google Scholar
-
Zhao, H. et al. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with Delta variant in TMPRSS2-expressed cells. Emerg. Microbes Infect. 11, 277–283 (2022).
Google Scholar
-
Yuan, S. et al. Pathogenicity, transmissibility, and fitness of SARS-CoV-2 Omicron in Syrian hamsters. Science 377, 428–433 (2022).
Google Scholar
-
Khan, K. et al. Omicron infection enhances Delta antibody immunity in vaccinated persons. Nature 607, 356–359 (2022).
Google Scholar
-
Lu, L. et al. Neutralization of severe acute respiratory syndrome coronavirus 2 Omicron variant by sera from BNT162b2 or CoronaVac vaccine recipients. Clin. Infect. Dis. 75, e822–e826 (2022).
Google Scholar
-
Ong, C. P. et al. Comparative analysis of SARS-CoV-2 Omicron BA.2.12.1 and BA.5.2 variants. J. Med. Virol. 95, e28326 (2023).
Google Scholar
-
Ye, Z. W. et al. Intranasal administration of a single dose of a candidate live attenuated vaccine derived from an NSP16-deficient SARS-CoV-2 strain confers sterilizing immunity in animals. Cell. Mol. Immunol. 19, 588–601 (2022).
Google Scholar
Funding
This study was supported by grants 2021YFC0866100 and 2023YFC3041600 from the National Key R&D Program of China; grants C7142-20GF and T11-709/21-N from Hong Kong Research Grants Council; emergency collaborative grant EKPG22-01 of Guangzhou Laboratory; and a grant from CVVT of Health@InnoHK (to D.-Y.J.) as well as grant 2023A1515011910 from Guangdong Natural Science Fund (to Z.‐W.Y.).
Author information
Authors and Affiliations
Contributions
S.Y., Z.-W.Y. and D.-Y.J. designed the experiments, analyzed the data, and wrote the manuscript. K.K.-W.T. provided key reagents and contributed to the conceptualization of the work. C.P.O., K.T. and P.-H.H.C. contributed to the design of the study and the writing of the manuscript, performed the experiments with the assistance of H.Z., T.-T.T., Y.X. and J.W., and analyzed the data. All authors reviewed, critiqued, and approved the text. C.P.O., K.T. and P.-H.H.C. contributed equally to this work.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethical approval
All protocols for animal experiments have been approved by the Committee on the Use of Live Animals in Teaching and Research (CULATR) of the University of Hong Kong (Reference code: CULATR 5754-21).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary information
REPORTING SUMMARY
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Reprints and Permissions
About this article
Cite this article
Ong, C.P., Tang, K., Cheung, PH.H. et al. Monovalent vaccination with inactivated SARS-CoV-2 BA.5 protects hamsters against Omicron but not non-Omicron variants.
npj Vaccines 8, 177 (2023). https://doi.org/10.1038/s41541-023-00776-x
-
Received: 19 May 2023
-
Accepted: 09 November 2023
-
Published: 20 November 2023
-
DOI: https://doi.org/10.1038/s41541-023-00776-x